

Зоофитовзаимодействия в регуляции средообразующих процессов
В системе ландшафтов пустыни Каракум, второй по величине в мире, преобладающее место принадлежит разнообразным типам песчаных массивов, различающимся своим рельефом, литологией, гидрологическими условиями и особенностями сложения и структуры биогеоценозов. Животные и растения здесь наряду с факторами абиотической среды оказываются мощными агентами средообразовательного процесса, влияющими на формирование рельефа поверхности, развитие ветроэрозионных процессов, почвенно-гидрологические условия (уровень грунтовых вод, водно-физические и химические свойства почв и миграцию солей в них), на структуру и плотность поверхности почвогрунтов, особенности микроклимата и в конечном итоге на структуру биогеоценозов и сам облик ландшафтов.
Взаимодействие организма с окружающей средой складывается в сложной и динамичной системе прямых и обратных (реципрокных) взаимосвязей, определяемой как зонально своеобразным уровнем дозирования факторов среды, так и диапазоном проявления биологических свойств организмов (например, пределами объема роющей деятельности грызунов, коррелированной с численностью и состоянием их популяций и т. п., или размерами фитогенного засоления почв галофильной растительностью, или, напротив, величиной мелиорирующего влияния некоторых видов деревьев и кустарников и др.).
В число подобных многообразных корреляций входят такие, изменение которых кардинально влияет на дальнейший ход процессов в биогеоценозе, определяет само направление функциональных связей и, следовательно, отражается в цикле развития всего биогеокомплекса. Эти связи следует относить к типу регуляторных. Они влияют на особенности структуры комплекса на разных стадиях биогеоценотических циклов. Определение момента и условий проявления биорегуляторных свойств взаимодействующих видов представляет собой сущность исследования процессов зоофитогенной регуляции в биогеоценозах как одного из главнейших механизмов средообразовательного процесса. Смысл и условия "биогенных поворотов" в цикле индивидуального развития зонального типа биогеоценоза служат важной основой для прогнозирования функциональной судьбы компонентов биогеокомплексов в условиях изменяющегося природного фона окружающей среды больших регионов.
Отдельные виды животных, растений и микроорганизмов могут резко различаться между собой по величине и значимости средообразующего эффекта, который они способны вызывать. Различия эти определяются прежде всего тремя условиями: 1) направлением воздействия вида (способно ли влияние жизнедеятельности вида вызывать единичную реакцию соседнего вида и комплекса в целом, или оно оказывается началом цепи процессов, которые к тому же могут стать групповыми процессами); 2) величиной охвата структурно-функциональных ярусов биогеоценоза (так, в случае развития зоофиторегуляторных процессов в одном-двух биогеоценотических ярусах изменения в структуре комплекса окажутся временными и затем через несколько лет нивелируются полностью, что, например, происходит при влиянии степных пеструшек на растительность полупустыни. В случае же вовлечения в процессы преобразования всей биогеоценотической толщи, всех ее ярусов, средообразующий эффект деятельности вида оказывается постоянным и входит в число ведущих факторов средообразования. Пример тому - роющая деятельность малого суслика в Прикаспии, сказывающаяся буквально на всех элементах природной среды и определяющая саму морфоструктуру биогеоценозов полупустыни); 3) степенью распространенности ценотически активного вида, что непосредственно определяет ареал и, следовательно, ландшафтную типичность его средообразующего влияния (виды массовые, доминанты, непременные участники комплекса обладают и наибольшей средообразующей значимостью).
Основной средообразующий эффект может возникать как результат взаимодействия, группы видов (при условии их сопряженного функционирования!), но оказывается нередко итогом преимущественного влияния вида, который в этом случае по праву может быть назван главным, или ведущим, средообразователем. Чаще всего вид - ведущий средообразователь - выступает в комплексе с группой сопутствующих видов ("виды-спутники"), смены группировок которых во времени определяются этапами кондиционирования среды под влиянием жизнедеятельности вида - главного ценозообразователя в разные возрастные периоды его жизни.
Участие животных в едином комплексе с растением - главным ценозообразователем и его спутниками может оказывать регулирующее влияние на развитие биогеоценотических процессов, на формирование структуры биогеоценоза, на смену и особенности течения стадий биогеоценотического цикла. Таким образом, изучение видоизменений средообразующего воздействия главного ценозообразователя под влиянием животных, т. е. зоорегуляции средообразовательных процессов, представляется существенным для характеристики биогеоценотического цикла в целом.
Для исследования процесса зоофитовзаимодействий в биогеоценозах южных песчаных пустынь Средней Азии были избраны зонально типичные для этих условий комплексы: 1) черного саксаула и песчанок, преимущественно полуденной и отчасти большой песчанки; 2) хвойника шишконосного и полуденной песчанки; 3) хвойника шишконосного, полуденной песчанки и черного саксаула и 4) черного саксаула, саксаулового усача и древесного паразитического гриба - пории пустынной (Poria desertorum).
Особенности черного саксаула как ценозообразователя
Черный саксаул (Haloxylon aphyllum (Minkw.) Iljin [H. ammodendron (С. А. Меу) Bge.]), "оджар" по-туркменски, - самый типичный вид дерева среднеазиатских пустынь; представляет собой мощный фактор ценозообразования как в глинистых, гипсовых пустынях, так и в песчаных. Естественная область распространения вида охватывает всю Среднюю Азию, южные районы центральной части Казахстана, Приаралье, плато Устюрт, пригодные для его произрастания участки закаспийских пустынь Мангышлака и полуострова Бузачи и выходит за пределы СССР на юго-запад, простираясь до северных берегов Персидского залива и Суэцкого перешейка (Леонтьев, 1954). Таким образом, примерно одна половина ареала черного саксаула оказывается расположенной в области, занятой пустынями южного типа, с субтропическим климатическим режимом и бесснежной или малоснежной зимой; другая - охватывает Туранскую подзону переходного характера, северные пустыни Закаспия и юга Казахстана в пределах умеренного климатического пояса с их экстремальными условиями - жарким летом и суровой, морозной и достаточно снежной зимой.
Ценозообразующая способность черного саксаула определяется относительным долголетием дерева, большим объемом его фитомассы, дающей много органического вещества и создающей "покровный эффект" - защитные условия и, следовательно, своеобразную среду для большого числа видов растений, животных, водорослей и микроорганизмов. Он влияет на водно-физические и химические свойства почвы, содействует миграции солей в толще почвогрунта и на его поверхности возникновению на почве корковых образований с фитогенным засолением и последующей сменой растительности в травянисто-кустарничковом ярусе (Базилевич, Чепурко, Родин, Мирошниченко, 1972). На полкроновом пространстве формируется специфический микроклимат, отличающийся относительной мезофильностью условий. Таким образом, влияние черного саксаула служит основой экологической дифференциации среды за счет создания особых микроусловий на разных, четко выделяемых парцеллах подкронового пространства.
Черный саксаул относится к жизненной форме одноствольных или немногоствольных афильных деревьев (Нечаева, Василевская, Антонова, 1973). В естественных древостоях встречается высотой 5-8 м, иногда 9 м и даже 12 м, но нередко при низком уровне грунтовых вод на плотном глинистом грунте или в условиях избыточного засоления в окрестностях шоров и соленых озер образует редкостойные заросли кустарникового облика до 1,4-2 м высоты. На кырах северо-западной части Туркмении на серо-бурых почвах встречается в виде особой экологической формы деревца, достигающего всего 1,2 м (Нечаева, 1951). Ствол начинает ветвиться дихотомически уже на высоте 50-70 см от поверхности земли. К 15-20-летнему возрасту рост дерева замедляется, и в это время ширина кроны оказывается равной ее высоте, представляя собой достаточно плотную темно-зеленую "шапку", низко нависающую над землей за счет опускания боковых сильно отклоняющихся ветвей. В Каракумах нередко встречается "плакучая форма", максимальная ширина кроны - 10-12 (14) м, средняя - 5-6 м.
Диаметр ствола спелых деревьев у поверхности почвы - 24-38 см. В исключительных случаях наблюдаются перестойные особи с очень толстым стволом, до 65 см, как, например, в черносаксауловом лесу у колодца Яр-Аджи, и даже 102 см, по замеру В. Л. Леонтьева (1954), в массиве у колодца Керт-Кую. Корневая система мощно развита, направлена в основном вертикально вниз и может достигать уровня относительно неглубоких грунтовых вод. Под загущенным или сомкнутым древостоем уровень грунтовых вод понижается из-за влияния сильной транспирации черносаксаулового леса (Леонтьев, 1954).
На закрепленных песках в долинообразных понижениях в Репетекском заповеднике (Восточные Каракумы), где грунтовые воды обнаруживаются на глубине около 7-10 м, корни черного саксаула распространяются в 4-6-метровой толще песка (Петров, 1933). В искусственных насаждениях в Бадхызе уровень грунтовых вод низкий, там корни черного саксаула прослеживались до глубины 7 м (С. Приходько и Н. Приходько, 1968), а в предгорной пустыне Карнабчуля (Самаркандская область), где уровень грунтовых вод очень низкий, корни 4-летнего черного саксаула в культуре проникают до глубины 14 м (Шамсутдинов, 1962, 1969).
Средний возраст деревьев в естественных древостоях Каракумов, лимитируемый, как выяснилось в результате исследований, комплексом экологических факторов, равен 40-43-45 годам и значительно меньше биологического возраста, достигающего 100 и более лет. Величина общей биомассы, а также соотношение надземной и подземной (корни) фитомассы и объем семенной продукции существенно изменяются в процессе индивидуального развития черного саксаула.
Вес надземной фитомассы взрослого дерева черного саксаула, по наблюдениям Р. К. Тогызаева, в Репетекском заповеднике достигает 152 кг. На приколодезных песках в Бадхызе 13-летнее дерево, по данным С. Я. Приходько, имело вес более 100 кг, при этом вес надземной части составлял 78,7 кг, а корней - 48,3 кг, т. е. в отношении 1:0,6 (Нечаева, Василевская, Антонова, 1973). Наибольший вес надземной фитомассы одного дерева достигает 500 кг (Леонтьев, 1954), запас древесины в среднем - 8-10 т/га, максимальный - до 60 т/га, урожай кормовой массы в естественных древостоях - около 10 ц/га, в искусственных насаждениях - 5-8-16 ц/га в воздушно-сухом состоянии (Шамсутдинов, 1969).
Плодоношение начинается в возрасте 4-6 лет (Леонтьев, 1946; Кокшарова, 1957; Пешковский, 1959), при этом в первый год плодоношения дерево дает в среднем 25 г семян (Кокшарова, 1957), во второй - до 500 г, в возрасте 7 лет - 900 г, а 10-12-летнее - 2160 г семян. Наиболее обильно плодоносят 12-35-40-летние деревья, когда с каждого можно получить от 0,2 до 10 кг семян (Нечаева и Приходько, 1966).
Урожай семян черного саксаула в культуре достаточно высок. Семена, собранные с 1 га насаждений, обеспечивают новый посев на площади 12-90 га (табл. 7).
| Возраст | Количество растений на 1 га |
Урожай семян, кг/га |
Коэффициент размножения |
| 6 | 910 | 62,0 | 12,0 |
| 7 | 910 | 305,0 | 61,0 |
| 12 | 660 | 313,0 | 62,5 |
| 13 | 660 | 452,0 | 90,5 |
Черный саксаул экологически пластичен и относительно устойчив к высоким летним температурам и зимним морозам при недостатке атмосферной и почвенной влаги. Особенно хорошо переносят низкие отрицательные температуры умеренного климатического пояса деревья из популяций северной половины ареала - устюртской и кызылкумской. Опыты акклиматизации черного саксаула в полупустыне Северного Прикаспия, на опытной станции ВНИАЛМИ ВАСХНИЛа в окрестностях станции Харабали Астраханской области и в районе оз. Баскунчак, показали способность некоторых особей этого вида не вымерзать даже в условиях очень суровых зим, при температуре воздуха до -30° С. Опыты по расселению и акклиматизации черного и зайсанского саксаулов к северу и западу за пределы их естественных ареалов в настоящее время проводятся в Астраханской, Волгоградской областях и Калмыцкой АССР.
Экологическая пластичность этого вида позволяет с успехом проводить искусственное расселение его по разным типам местообитаний внутри естественного (исходного) ареала и восстанавливать сведенные массивы, что представляется наиболее перспективным с точки зрения принципов биогеоценотической мелиорации пустынь, вполне технически осуществимо и может быстро дать весьма существенный экономический эффект за счет резкого повышения продуктивности пустынных пастбищ.
Типичные местообитания черного саксаула в песчаных пустынях - это долинообразные понижения с закрепленной поверхностью среди массивов грядовых и барханно-грядовых песков (Восточные Каракумы), днища и склоны котловин в крупнобугристых песках с присутствием гипса и с относительно неглубокими грунтовыми водами (4-7-10 м). При этом в зависимости от их уровня может изменяться характер древостоя, его густота, ход возобновления, состав и обилие растений-спутников и животное население.
При близко расположенных к поверхности и не слишком засоленных грунтовых водах возникают загущенные и сомкнутые древостой с числом стволов до 400-560 на гектар, представляющие собой настоящие леса в пустыне. При этом возобновление в таких насаждениях слабое и усиливается лишь в перестойных участках, где имеются осветленные места под усохшими ветвями и на месте выпавших особей. Среди сопутствующих видов нижнего яруса появляются в виде зарослей верблюжья колючка (Alchagi persarum Boiss et Buhse) и акбаш-карелиния (Karelinia caspia (Pall.) Less.), служащие индикатором близости грунтовых вод (например, черносаксаульники вдоль трассы первой очереди Каракумского канала, ныне почти полностью сведенные). При глубине грунтовых вод 7-12 м черносаксаульники более редкостойны (190- 260 взрослых особей на 1 га) и нередко сочетаются с эфедрой - хвойником шишконосным, особенно обильным на гипсоносных песках (обычно на буграх), и с несколькими видами кандымов (Callygonum). Эфедра может служить надежным индикатором значительных количеств гипса в почвогрунтах, который нередко образует массивные, порой причудливые друзы из крупных кристаллов, что столь типично для репетекских песков.
В составе растений-спутников эуксерофиты и ксеромезофиты, эфемеры и эфемероиды, злаки и однолетние солянки (Londesia eriantha Fisch. et Mey). При еще более глубоких грунтовых водах на полузакрепленных песках черный саксаул растет в сочетании с белым саксаулом (Haloxylon persicum), древовидным и другим кандымами, черкезом Рихтера (Salsola richteri) и чогоном (Aelenia subaphilla) в Центральных, Восточных и Заунгузских Каракумах. В числе сопутствующих видов на определенных этапах ценотического цикла добавляются настоящие псаммофиты - гораниновия (Horaninovia ulicinia Fisch. et Mey), гелиотропы, смирновия и др. Растет черный саксаул и на шлейфах песков, выходящих на глинистые равнины и понижения вокруг шоров и соленых озер (например, в Бадхызе), где характер древостоя и высота деревьев в зависимости от условий произрастания могут быть очень различными.
В пустынях с плотным грунтом (глинистых, гипсовых и суглинистых с серыми и серо-бурыми слабо сформированными почвами) черный саксаул также образует древостой лесного типа и редколесья. При этом может произрастать на территориях с очень глубоким залеганием грунтовых вод (глубже 20-30 м), как, например, на плато Южный Устюрт, где естественные древостой в настоящее время сильно нарушены (Викторов, 1971), и в культуре на Карнабчуле в Узбекистане (Шамсутдинов, 1969, 1971, 1973; Ибрагимов, 1975). Способен обитать саксаул и в условиях сильно расчлененного рельефа, на крутых склонах оврагов - саев, на каменистых и насыщенных щебнем пятнах делювия на обрывах - чинках плато. В горы поднимается до 500-800 м (по наблюдениям Г. М. Проскуряковой и автора, в хребте Большой Балхан).
Влажность почвогрунта под черным саксаулом. Развитые деревья саксаула, создающие сильное затенение, плотную засоленную поверхностную корку на почве подкронового пространства и нередко достигающие своей мощной корневой системой горизонта грунтовых вод, существенно влияют на распределение и динамику почвенной влаги, образуя мозаику почвенно-гидрологических условий. Последнее можно легко обнаружить при раскопке песка уже до глубины 0,5-1,0 м. Определение влажности почвогрунта проводилось в спелом эфедровом черносаксаульнике, растущем в долинообразном понижении между барханными грядами на территории Репетекского заповедника (Восточные Каракумы). Здесь, в Северной долине с несформированными песчаными почвами, под закрепленными илаком (Carex physodes) кустарниками и злаково-солянковыми группировками подкроновой растительности песками, в качестве модели было выбрано спелое дерево в возрасте около 30 лет, высотой 3,8 м и диаметром кроны 3,6X6,0 м (с характерной асимметрией). Диаметр подкронового пространства вместе с кольцом бордюрной растительности из однолетних солянок с участием злаков и эфемерового разнотравья (4,3X6,5 м) оказался больше диаметра кроны, что объясняется ее частичным усыханием и расширением кольца бордюрной растительности в теневой зоне дерева, выходящей за пределы непосредственно проекции кроны.
Пробы почвогрунта брались с помощью почвенного бура в 7 точках, 5 из которых находились на подкроновом пространстве: 1) среди злаково-солянковых группировок бордюрной растительности с северной стороны от ствола саксаула, на границе с поляной, занятой дерниной илака; 2) на оголенном пятне фитогенного засоления под кроной к северу от дерева; 3) на пятне подкронового засоления с южной стороны от ствола дерева на затененном его участке; 4) на пятне засоления в одном метре от предыдущей точки, но на осветленном участке под усохшей веткой и 5) на бордюрном кольце солянково-злаковой растительности с южной стороны от ствола. Две другие точки представляли собой контроль и находились на илаковых полянах к северу и к югу от дерева в 2 м от края подкронового пятна. Влажность почвогрунта определялась в начале (6 июня) и в конце (10 августа) лета.
Исследования показали, что как в начале лета, так и в его конце почвогрунт на подкроновом пространстве черного саксаула имеет большую влажность, чем соответствующие горизонты почвы под микрогруппировками бордюрной растительности и тем более под песчано-осоковой ассоциацией на полянах (в первой декаде июня в горизонте 0-5 см под кроной на пятне фитогенного засоления - 1,1-1,2%, под солянково-злаковым бордюрным кольцом - 0,4-0,5%, на илаковой поляне (контроль) - 0,2-0,4%). На протяжении лета показатели влажности в верхних горизонтах почвогрунта (0-20-30 см) на подкроновом пространстве заметно снижаются: в горизонте 0-5 см - от 1,2% до 0,6%, в горизонте 5-20 см - от 3,0% до 0,9-1,8%.
Особенно иссушается приповерхностный горизонт почвы (0-5 см) в течение лета, и показатели его влажности на оголенном пятне засоления под кроной и под бордюдной растительностью (0,6%) в августе становятся одинаковыми до глубины 20 см, приближаясь к показателям влажности соответствующих горизонтов на илаковых полянах (0,4-0,3%). Однако в горизонтах почвогрунта от 20-40 см и ниже в корнеобитаемой зоне под проекцией кроны черного саксаула влажность почвы в конце лета может втрое и более (1,2-1,8%) превышать ту, что наблюдается на илаковых полянах на глубине более 40 см (0,3-0,5%).
Учитывая сказанное, можно констатировать, что взрослое дерево черного саксаула, обладающее глубокой корневой системой (до 10-14 м) и активно транспирирующее, влияет на распределение почвенной влаги, содействуя ее сохранению под уплотненной поверхностной коркой (толщиной 0,8-1,3 см) фитогенного, преимущественно сульфатного, засоления, ограничивающего непосредственное испарение с поверхности почвы. Этому способствуют также затенение кроной и более низкие температуры поверхности на подкроновом пространстве (см. рис. 5). Кроме того, в корнеобитаемой зоне средневозрастного и спелого дерева в весенне-летний период происходит подтягивание почвенной влаги из более глубоких горизонтов почвогрунта.
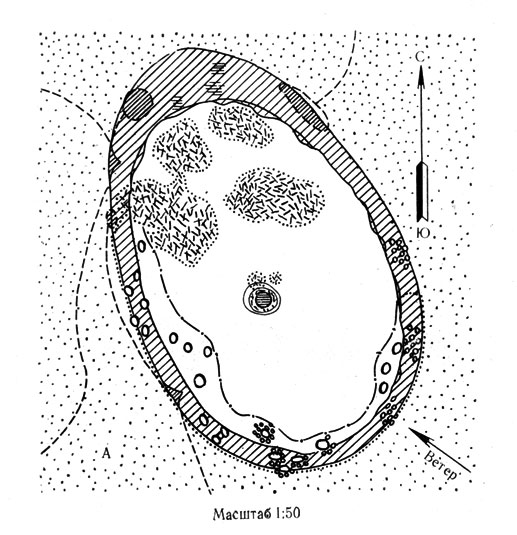
Рис. 5А. Схема подкронового пространства черного саксаула и солянки (парцеллы подпокровного пространства) - биогеоценотическая система 'вложенных структур' (южная пустыня, Восточные Каракумы)
В связи с этим распределение почвенной влаги по профилю вертикального разреза почвогрунта от илаковой поляны через подкроновое пространство спелого саксаула имеет вид "купола", образованного зоной повышенной влажности непосредственно под деревом вдоль оси корня со "склонами", довольно круто спадающими под бордюрным кольцом растений и еще более резко при переходе от бордюра к илаковым полянам. В течение лета этот "купол" постепенно оседает, сохраняя между тем свои общие очертания с вершиной, находящейся в центральной части подкронового пространства.
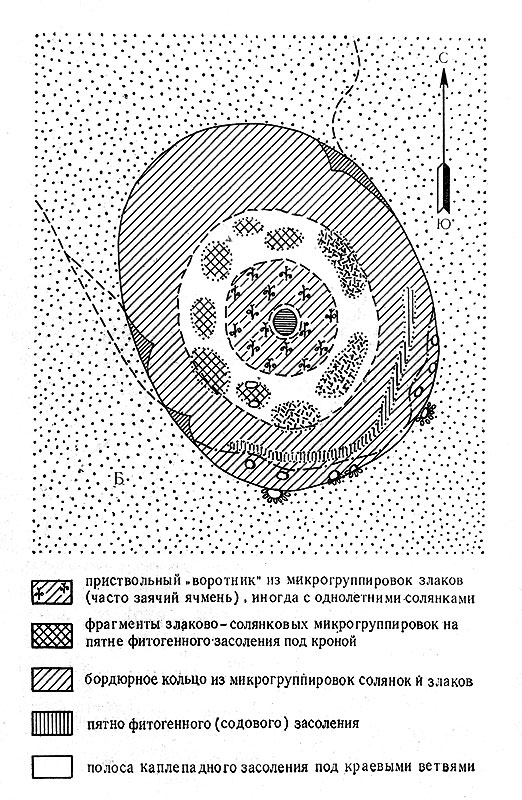
Рис. 5Б. Схема подкронового пространства черного саксаула и солянки (парцеллы подпокровного пространства) - биогеоценотическая система 'воротниковых структур', туранский вариант системы 'вложенных структур' развит в районах с повышенным атмосферным увлажнением (предгорная пустыня, Карнабчуль)
Таким образом, черный саксаул оказывается экологическим агентом, создающим разнообразие почвенно-гидрологических условий и формирующим мозаичное распределение "гнезд влаги" в пустыне, длительно сохраняющихся в летний сезон под коркой засоленного пятна оголенного подкронового пространства, которое оказывается, таким образом, не только пескозакрепителем, но и влагосохраняющим природным устройством (Залетаев, Зубкова, 1968) и играет роль сезонно действующего гидротермического рефугиума для ряда видов позвоночных и беспозвоночных животных. Свидетельством этого служит сезонная смена стаций, наблюдаемая у некоторого количества полуденных песчанок в Каракумах. Если зимой большинство их обитает в буграх эфедры шишконосной, где суше, а температурная среда более стабильна, то в летний сезон часть нор под эфедрой пустеет в связи с переселением их обитателей под соседние деревья черного саксаула. Здесь, в норах на периферийной части засоленного подкронового пятна, создаются более комфортные для зверьков микроклиматические условия, определяемые свойствами черного саксаула как ценозообразователя, создающего в зоне своего влияния особую динамику тепла и влаги и формирующего собственные группировки растений и животных-спутников.
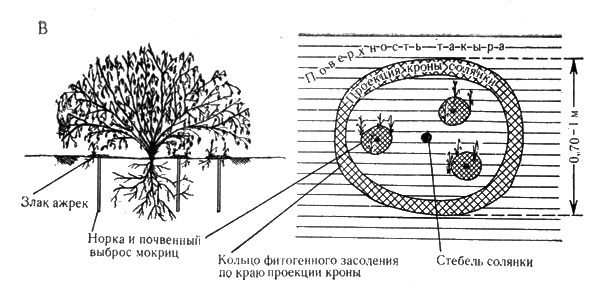
Рис. 5В. Схема подкронового пространства черного саксаула и солянки (парцеллы подпокровного пространства) - система 'вложенных структур', развитая в ажреково-солянковом комплексе (южная такыровая пустыня, долина р. Сумбар)
Роющая деятельность песчанок - как фактор, содействующий возобновлению черного саксаула и кустарников-псаммофитов, - исследовалась в разных типах древесно-кустарниковых биогеоценозов песчаных пустынь. Наблюдения показали заметную концентрацию подроста и молодых (2-4-летних) деревцов черного саксаула, и подроста разных видов кандымов (Calligonum setosum, С. turkestanicum, С. caput medusae, С. microcarpum, С. arborescens и других), а также черкезов (Salsola richteri, S. paletzkiana) на колониях песчанок и у выходов из нор, расположенных небольшими группами.
Помимо фиксации многочисленных случаев активного возобновления черного саксаула и кустарников-псаммофитов в зоне нор песчанок (см. табл. 8) были проведены учеты подроста на пробных площадях в кандымово-илаковом белосаксаульнике, в том числе на участке с длительно существующей колонией больших песчанок котловино-лощинного типа и в эфедровом черносаксаульнике, типичном для условий Восточных Каракумов. На этих площадях было учтено количество и размещение нор большой и полуденной песчанок, число и распределение подроста саксаулов обоих видов и кустарников и степень приуроченности подроста к зонам нор, что отражалось процентным соотношением подроста у нор и вне зоны нор.
| Виды деревьев и кустарников-псаммофитов |
Количество деревьев и кустарников, шт/га |
Большая песчанка | Полуденная песчанка | Общее число, шт/га |
||
| число шт/га |
% | число шт/га |
% | |||
| Haloxylon persicum | 80 | 410 | 38,0 | 50 | 18,5 | 460 |
| Haloxylon aphyllum | 90 | 110 | 10,5 | 50 | 18,5 | 160 |
| Calligonum setosum | 25 | 35 | 3,5 | 40 | 14,8 | 75 |
| Calligonum turkestanicum | 35 | 160 | 15,0 | 25 | 9,2 | 185 |
| Ephedra strobilacea | 5 | 0 | * | 55 | 20,5 | 55 |
| Aellenia subaphylla | 10 | 20 | 2,0 | 0 | - | 20 |
| Salsola paletzkiana | 5 | 0 | * | 0 | - | 0 |
| Поляны с Carex physodes* | - | 330 | 31,0 | 50 | 18,5 | 380 |
| Всего | 250 | 1065 | 100% | 270 | 100% | 1335 |
* (Число выходов из нор, расположенных вне подкроновых пространств.)
Пробная площадь (0,2 га) была заложена в окрестностях Казахаула за пределами Репетекского заповедника в редкостойном кандымово-илаковом белосаксаульнике с участием черного саксаула (состав насаждения представлен в табл. 8). Общее число стволов деревьев и число кустов - 250 шт/га, в том числе белого саксаула - 80 шт/га, из них: взрослых особей - 65 шт/га, подроста - 15 шт/га (18,8%); черного саксаула - 90 шт/га: взрослых - 40 шт/га и подроста - 50 шт/га (55,5%); кандыма щетинистого (Catligonutn setosum) - 25 шт/га: взрослых - 5 шт/га и подроста - 20 шт/га (80%); кандыма туркестанского (Calligonum turkestanicum) - 35 шт/га: взрослых особей - 10 шт/га и подроста - 25 шт/га (71,4%).
Кроме того, на этой площадке произрастали отдельные единичные кусты черкеза, чогона (Aellenia subaphylla) и борджока (Ephera strobilacea), последние образовали довольно развитый песчаный бугор, заселенный исключительно полуденными песчанками. На всей площадке 267 выходов из нор песчанок: большой песчанки - 213, или 1065 нор/га, полуденной песчанки - 54, или 270 нор/га. Суммарная насыщенность норами поверхности песка в кандымово-илаковом белосаксаульнике с большой колонией песчанок достигает, таким образом, 1335 нор/га.
Учет размещения нор и топографическая съемка пробной площадки показали, что из общего количества подроста (110 шт/га деревьев и кустарников-псаммофитов) непосредственно в зоне нор произрастает 71 шт/га (64,8%).