

Плодовитость
Беременность лосих продолжается 225-237 дней (Кнорре, 1959). Ко времени отела самки отгоняют своих прошлогодних телят и те вынуждены вести самостоятельную бродячую жизнь. Новорожденных лосят (рис. XVI, XVII) мы наблюдали начиная с 24 апреля (1964 г.), но в литературе для Лужского района упоминается еще более ранняя дата - 21 апреля 1959 г. (Фетисов, 1959). Массовый отел приурочен к первой половине мая. Часть коров, главным образом молодых, телится во второй половине мая - начале июня. Например, 11 июня 1964 г. мы встретили молодую лосиху с двумя лосятами, один из которых еще неуверенно передвигался, второй же не отставал от матери.

Рис. XVI. Новорожденный лосенок. Фото Ю.Б. Пукинского

Рис. XVII. Первые шаги новорожденного лосенка. Фото Ю.Б. Пукинского
Сравнение показывает, что отел в Ленинградской обл. происходит в те же сроки, что и в Дарвинском заповеднике (Калецкая, 1961), тогда как в Карелии, Лапландском заповеднике и в скандинавских странах массовый отел имеет место в конце мая-начале июня, причем в Норвегии и Лапландии новорожденные лосята могут появляться даже в июле (Семенов-Тян-Шанский, 1948; Лебедева, 1968). На Верхней Печоре лоси телятся с первой декады мая по первую декаду июня (Кнорре, 1959).
Судя по наблюдениям в природе, к сожалению недостаточно многочисленным, коровы с одним теленком встречаются в два с лишним раза чаще, чем с двумя. Однако в отдельные годы и особенно по сезонам в течение одного года эта картина существенно меняется (табл. 8 и 9). Как видно из данных табл. 8, средний процент двоен в отдельные годы может уменьшаться до 22,5-23,4, как то было в 1963 ,и 1966 гг.
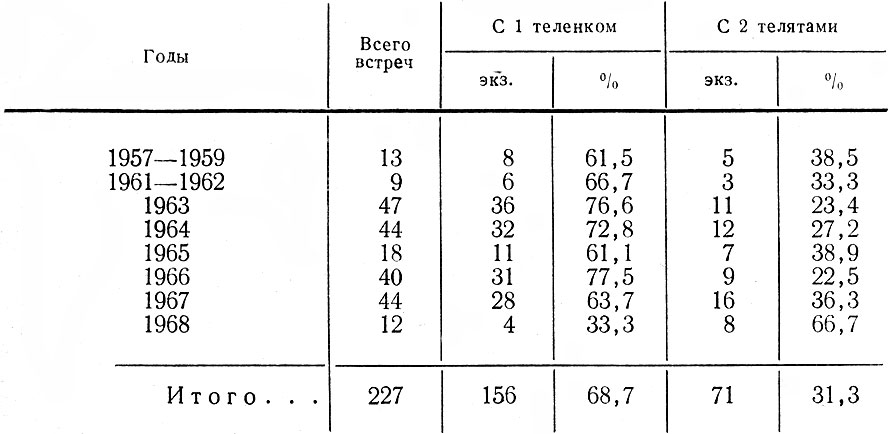
Таблица 8. Встречи коров с одним и двумя телятами по годам
В мае-июне двойни составляли 41,6% встреч новорожденных телят. Заметим, что на Кольском п-ове они отмечены в 43% случаев (Семенов-Тян-Шанский, 1948), в Карелии - в 31,0 (Троицкий, 1972), в Дарвинском заповеднике - в 42,7 (Калецкая, 1961), в Завидовском охотничьем хозяйстве под Москвой - 43,5% (Макарова, 1969), в Печоро-Илычеком заповеднике в разные годы процент двоен колебался от 29 до 53 (Кнорре, 1959). Далее, как свидетельствуют данные табл. 9, ко времени окончания гона количество двоен уменьшается более чем на 19%, повидимому, за счет гибели одного из телят. Но основной отход молодых в парных пометах происходит зимой, так что во второй ее половине встречаются коровы преимущественно с одним теленком (82,1-%). В связи с этим интересно привести данные Р. Реш (Resche, 1968) о смертности телят на Аляске в весеннеосениий период. В этой небольшой изолированной популяции, насчитывающей около 300 голов, было помечено 59 новорожденных телят. 56% из них погибли в первые 5 месяцев, преимущественно в первые две недели. При этом среди двоен в течение одного месяца погиб 31 %, среди одиночек - только 9%.
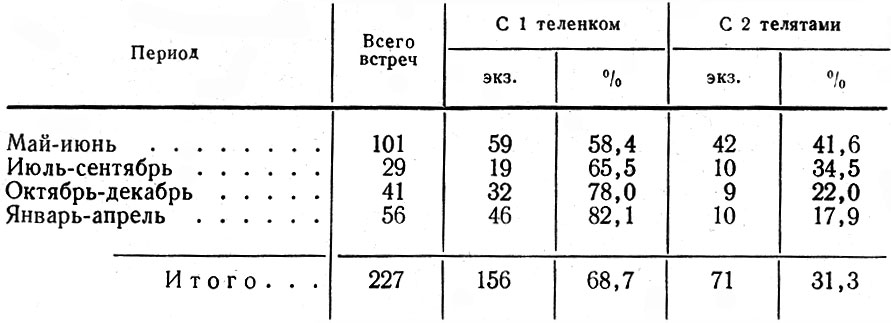
Таблица 9. Встречи коров с одним и двумя телятами по сезонам в 1957-1968 гг.
Если сопоставить приведенные выше данные визуальных наблюдений со сведениями в лицензиях, сданных в Госохотинспекцию за 5 лет, то обнаружится значительное между ними расхождение (табл. 10). Согласно лицензиям, среди добытых 3364 стельных лосих с одним эмбрионом было 2679 экз. (79,6%), с двумя - всего 681 (20,3%), а у четырех самок (менее 0,1%) обнаружено даже по три эмбриона. Иными словами, по этим данным, число близнецов оказывается вдвое меньшим, чем фактически наблюдается в природе в мае-июне. Столь явное несоответствие можно объяснить лишь недостаточной точностью сведений, поступающих от охотников. Они нередко определяют число эмбрионов, ограничиваясь самым поверхностным осмотром матки, (без вскрытия ее. Такой способ неизбежно должен давать заниженные результаты, особенно если учесть, что в первые месяцы охоты на лосей эмбрионы могут быть еще очень слабо развитыми (имея длину менее 5 см) и поэтому обнаруживаются только при тщательном осмотре матки*. Этим же обстоятельством объясняются явно завышенные сведения о яловости отстрелянных лосих. Тем не менее, процент яловых половозрелых коров все же очень велик.
*(К аналогичному выводу пришел Г. А. Троицкий (1972) в отношении Карелии.)
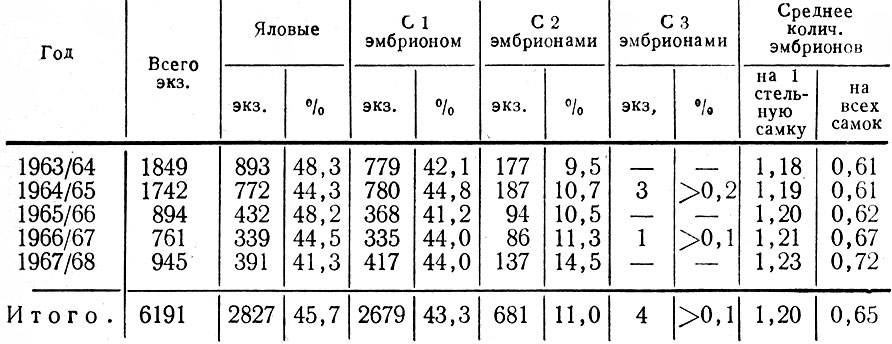
Таблица 10. Плодовитость лосих по данным лицензионного отстрела 1963 - 1968 гг.
За последние 6 лет в Ленинградской обл. наблюдается неуклонное сокращение численности лося, после того как она достигла своего максимума в 1962 г. Ухудшение кормовой базы, истощение животных в районах с чрезмерно высокой плотностью популяции приводит к снижению их плодовитости, в первую очередь за счет повышения яловости, которой сопутствует усиленная смертность и сокращение числа парных пометов.
Падение плодовитости популяций лося к концу 50-х - началу 60-х годов нашего столетия наметилось во многих областях европейской части СССР (Киселева, Приклонский, Теплов, 1965). В Ленинградской обл. наибольшее количество яловых коров (50%) было отмечено в 1965 г. в Сосновеком лесоохотничьем хозяйстве, где на 1000 га насчитывалось до 53 лосей (Ким, 1967 а). Сходная картина отмечалась и для ряда популяций североамериканского лося. По данным Д. Пимлота (Pimlott, 1961), в районах с высокой плотностью населения самки по второму году не участвуют в размножении, а взрослые имеют только 3% двоен. В местах с низкой плотностью населения 67% полуторагодовалых самок беременны, а количество лосих с парными эмбрионами достигает 41 %.
По наблюдениям Е. П. Кнорре (1959), большинство молодых лосих приносят потомство только по достижении веса не менее 290 кг; плохо упитанные старые самки также остаются яловыми, хотя участвуют в гоне. Вес ленинградских лосей значительно меньше, чем печорских, но указанная закономерность прослеживается не менее четко. Мы, к сожалению, не можем точно отнести добытых при массовых отстрелах лосей к той или иной возрастной группе, так как охотники обычно определяют возраст самым (приблизительным образом. С наибольшей точностью, повидимому, можно установить возраст молодых (2-3-годовалых) животных и старше 10 лет. Вес лосих в возрасте 2,5-3,5 лет не превышает 250 кг. Яловость в этой группе достигает 65% и на одну стельную самку приходится 1,2 теленка. В последующих возрастных группах наименьший процент яловости (31), при показателе плодовитости - 1,2, отмечается у коров несом около 300 кг. У самок старше 10 лет яловость снова возрастает до 42,8%., Однако, как указывает Ю. П. Язан (1964), в природе, практически не встречается лосей, не способных к размножению вследствие глубокой старости, так как они просто не доживают до такого возраста и гибнут от тех или иных причин. Наибольшее количество двоен (до 25%) отмечается у крупных коров в возрасте 6-9 лет. Для сравнения укажем, что в Финляндии в начале 60-х годов плодовитость лосей была еще очень высокой. Исследование самок, добытых в 1963 г., показало, что среди молодых животных яловость не превышала 20%, старые же самки были все беременными и 80% из них имели двойни (Rajakoski, Koivisto, 1966).
Особого внимания заслуживает наличие самок с тремя эмбрионами. Известно, что тройни у лосей наблюдаются очень редко. Среди 3364 беременных самок, добытых в Ленинградской обл. за 5 лет, только 4 имели по 3 эмбриона. О нескольких случаях подобного многоплодия, известных от егерей и местных охотников, сообщают С. А. Бутурлин (1890), Е. П. Кнорре (1959) и Ю. П. Язан (1964). Однако только один факт описан специалистами (Павлов и Язан, 1967): в 1965 г. в Кировской обл. была добыта хорошо упитанная лосиха в возрасте 6,5-7,5 лет, имевшая 3 эмбриона, два из которых, самец и самка, весили 280 и 290 г, а третий (самец) всего 148 г. По заключению авторов, не была исключена возможность резорбции последнего. Три эмбриона были обнаружены два раза в Тамбовской обл. (Херувимов, 1969) и однажды в Швеции (Lonberg, цит. по Гептнеру и др., 1961). В связи со сказанным вызывает недоумение утверждение Р. А. Девишева (1967), что в Саратовской обл. в 1963/64 гг. 32,6% стельных лосих имели по 3 эмбриона, а в 1967-1969 гг. этот процент упал до 0,2. Ясно, что данное заключение основано на недоразумении, вызванном совершенно некритическим использованием материала лицензий. Стремление автора объяснить упомянутый феномен оптимальными условиями обитания лосей в Саратовской обл., сравнительно с другими частями ареала, не может быть признано состоятельным. Нельзя также не отметить исключительную небрежность Девишева в обработке и изложении данных. В своих публикациях (1967, 1970 а, б) он относит цитированные сведения то к 1963/64, то к 1962/63 годам, допускает в подсчетах грубые арифметические ошибки и т. д.
Плодовитость популяции - один из главных факторов, определяющих ее прирост. Однако при установлении фактического увеличения стада необходимо учитывать не только число новорожденных, но также гибель сеголетков, молодых и взрослых животных в течение года. В таежных районах плодовитость лося в основном определяется состоянием кормовой базы и метеорологическими условиями снежного периода. После продолжительных и снежных зим она резко падает, телята родятся мелкими, слабыми, и смертность среди них на протяжении года бывает выше обычной (Теплой, 1948; Кнорре, 1959). X. И. Линг (1969) считает условия зимы одним из важнейших факторов, определяющих структуру и прирост популяции лося в Эстонской ССР. Е. Райакоски и И. Койвисто (1970) видят одну из главных причин падения плодовитости финской популяции лосей в перепромысле, вследствие которого уменьшается процент взрослых животных и происходит непрерывное омоложение стада.
По данным регистрации встреч животных за последние 11 лет, новорожденные лосята с конца апреля по начало июня в среднем составляли 28,8% всех учтенных особей. К концу же снежного периода (в марте-апреле) годовалых лосят было только 14,6%. Следовательно, за 10-11 месяцев погибло около 50% телят. Ко времени достижения ими половой зрелости гибнет еще не менее 20%. Таким образом, отход молодняка к следующей осени достигает 70-75% (Херувимов, 1969; Приклонекий
Нам, к сожалению, известно только 12 случаев гибели новорожденных телят, что составляет 3% от всех установленных фактов гибели лосей в области. 3 теленка были убиты медведем, из них 2 (новорожденные) - в мае, 1 - в сентябре; 4 - погибли от волков в мае и июне; 4 новорожденных лосенка были найдены мертвыми без видимых повреждений; 1 - утонул, попав в воронку, заполненную водой.
Заканчивая рассмотрение вопросов, связанных с размножением изучаемой популяции, необходимо остановиться на имеющихся у нас данных об эмбриональном развитии лося в связи с малочисленностью такого рода сведений в литературе.
Нередко определение веса и размеров эмбрионов, добытых во время отстрела лосих, помогает правильно судить о характере гона и колебании его сроков. К сожалению, эмбриональное развитие лося изучено очень слабо. В частности, нет данных, позволяющих по внешнему виду, размерам и весу плода определить его возраст, а следовательно, и время оплодотворения самки. Мы присоединяемся к справедливой критике М. В. Кожуховым (1961) тех авторов (Лихачев, 1955, и др.), которые определяют время развития эмбрионов, руководствуясь одними общими сведениями о начале, разгаре и окончании гона, а не точными датами покрытия лосих. Не случайно, пытаясь определить возраст эмбрионов по схеме В. Д. Херувимова (1969), мы получаем во многих случаях явное завышение его и соответственно сдвиг сроков гона более чем на месяц. По-видимому, более точно описал этапы развития плода лося А. К. Петров (1964), хотя он не указывает происхождение своего материала. Вопросам внутриутробного развития лося, а также прижизненной диагностике беременности у лосих Верхней Печоры посвящены весьма обстоятельные работы М. В. Кожухов а (1959, 1961) и Л. И. Иржака (1967).
В течение 1965-1968 гг. нами было исследовано 68 эмбрионов, полученных из разных районов области при лицензионном отстреле лосей. Среди коров, от которых были взяты эмбрионы, преобладали 3-4-летние (57%) и 5- 6-летние (33%). 8 из них имели двоен.
Несмотря на ограниченность материала, характеризующего только первые 3-4 месяца эмбрионального развития, он ясно свидетельствует о том, что оплодотворение отдельных лосих происходит в течение всего периода гона, и розница но времени развития плода у них может достигать 2-2,5 месяца. В результате в ноябре вес зародышей колеблется от 3,5 до 264 г, в декабре - от 0,9 до 385 г, в январе - от 401 до 1590 г (табл. 11).
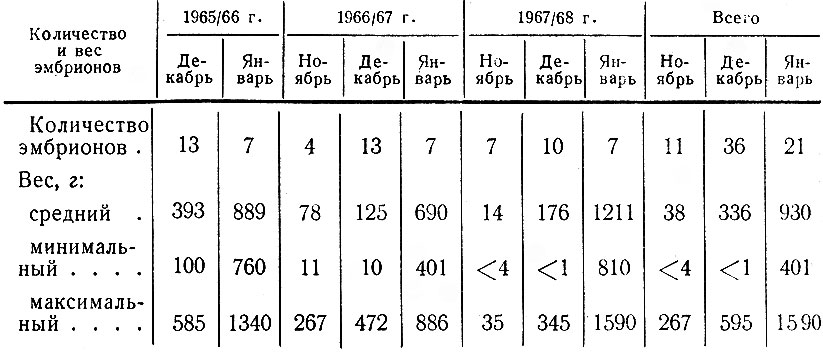
Таблица 11. Изменчивость веса эмбрионов по месяцам
М. В. Кожухов (1961) замечает, что столь существенная разница в весе эмбрионов у лосих, добытых в один и тот же месяц, определяется рядом факторов. Кроме погодных условий, нередко обуславливающих интенсивность гона, на время наступления течки у лосих влияет их возраст и упитанность. Известно, что молодые, болевшие и плохо упитанные коровы приходят в охоту значительно позже хорошо упитанных взрослых животных. По наблюдениям Е. П. Кнорре (1959), у одних и тех же самок в течение гона может быть несколько течек, повторяющихся через 19-20 дней. Здоровые коровы обычно оплодотворяются при наступлении первой течки, слабые, истощенные - при второй или даже третьей.

Рмс. 7. Эмбрионы лося разного возраста. а - вес 1,0 г, длина 37.м; б - 3,5 г, 65 мм; в - 14 г, 92 мм, самка;. г - 120 г, 168 мм, самка; д - 590 г, 300 мм, самка, возраст 3 месяца
Вес эмбрионов-двоен бывает несколько меньше, чем одиночек на той же стадии развития. Прямой же зависимости веса и размеров эмбриона от веса и состояния коровы на первых стадиях развития не наблюдается. Однако вес новорожденных телят, по данным указанных авторов, может колебаться в пределах 6,9-16 кг.

Таблица 12. Изменение размеров (см) эмбрионов с увиличением их веса (г)
Не располагая точными данными о времени покрытия лосих и возрасте эмбрионов, при описании мы руководствовались их весом и размерами, а также временем добычи коров (табл. 12). В таблице мы разместили эмбрионы в порядке, характеризующем их вес и размеры на разных стадиях развития. Самый маленький зародыш (рис. 7, а), возраст которого, согласно
А. К. Петрову (1964), несколько больше месяца, был обнаружен у коровы 5 лет, добытой 9 декабря 1967 г. в Тосненском районе. Вес эмбриона едва достигал 1 г, длина равнялась 3,7 см. Однако у него уже была хорошо развита голова с очень короткой лицевой частью, выраженным ротовым отверстием, глазными яблоками и ушными раковинами, передние и задние ноги, причем на передних были заметны два пальца. Сквозь тонкие покровы просвечивали кости черепа, туловища и конечностей. Пол определить не удалось, так как наружные половые органы были не выражены. Несколько больший эмбрион (3,5 г, 6,5 см) был извлечен из коровы 5 лет в Выборгском районе 19 ноября 1967 г. (рис. 7, б). В отличие от первого эмбриона у него были более отчетливо выражены ротовое отверстие, ноздри, уши, хвост. На всех конечностях, очень тонких, развито по четыре пальца. Пол, как и в первом случае, определить не удалось. У эмбрионов весом более 10 г определение пола уже не составляет труда (рис. 7, в).

Рис. 8. Эмбрион лося в возрасте 4 месяцев, вес 1590 г, самец
До 2-3 месяцев развития плод, достигший веса более 100 г, кроме короткого хвоста и сравнительно длинных ушей, не имеет еще видовых признаков (рис. 7, г). Ноги у этих эмбрионов тонкие, относительно короткие, голова большая, круглая, морда не вытянута. Эмбрионы, добытые в декабре-январе на четвертом месяце беременности коров, нередко превышают 400 г и начинают приобретать характерные видовые черты: удлиненную голову с вытянутой лицевой частью, длинные ноги с относительно большими копытами, хорошо выраженную холку (рис. 7, д). Когда вес эмбрионов достигает 700-800 г, у них начинают появляться первые волоски, сначала на нижней губе, затем на верхней, спинке носа и веках, у более старших- пигментированы копыта (рис. 8). Иногда заметны половые различия в телосложении: у бычков массивное и короткое туловище и толще шея, чем у телочек. Однако заметной разницы в весе одиночных эмбрионов разного пола на одной стадии развития не отмечается, В трех известных случаях разнополых двоен самцы были несколько тяжелее самок.
Сравнивая вес и размеры эмбрионов в исследуемом районе с аналогичными данными по Печоро-Илычскому заповеднику (Кожухов, 1961), мы находим, что в нашей области эти показатели для одних и тех же месяцев или сходны или несколько выше, что объясняется более ранним наступлением гона у нашей популяции лосей. Однако, с другой стороны, в Ленинградской обл. наблюдаются значительно более резкие колебания веса эмбрионов, добытых в один и тот же месяц. Это явление, по-видимому, объясняется тем, что в 1965-1968 гг. вследствие высокой численности лосей и истощения кормовой базы животные ко времени гона не нагуливали необходимого веса и часть из них ослабевала. В результате некоторые лосихи, особенно молодые, по упоминавшимся выше причинам, оплодотворялись позднее. Часть оставалась яловыми, тогда как в Печоро-Илычском заповеднике яловость взрослых лосих - явление крайне редкое (Кожухов, 1961).