

Морфоструктура биогеоценоза эфедровых черносаксаульников восточных каракумов
В системе ландшафтов южных пустынь биогеоценоз эфедровых черносаксаульников представляет собой древнюю устойчивую экосистему, что доказывается данными пыльцевого анализа о совместном распространении саксаулов и эфедры, относящимися к третичному времени. Развитый биологический комплекс черносаксаулового леса состоит из сочетания разновозрастных деревьев саксаула (в типе преобладают 20-25-летние) с обязательно сопутствующим им сложным подкроновым фитозоопочвенным образованием, называемым далее "подкроновой салфеткой", и кустов эфедры, около 2/3 которых растут на сформированных "эфедрогенных буграх". Эта система дополняется участием кандымов в различных сочетаниях видов и реже - белого саксаула.
Указанные модификации, даже в случае небольшого преобладания черного саксаула над сопутствующими породами, не изменяют данного географического типа биогеоценоза, который можно обозначить как систему: черный саксаул с "подкроновой салфеткой" - сеть ее солянково-злаковых бордюров (вокруг подкроновых салфеток) - песчанки - муравьи + эфедра (на буграх) и песчанки. В дополнительную характеристику биоком-плекса входят саксауловый усач, чернотелки, златки, саксауловый сеноед, пауки, скотоцерки, малые бормотушки, рыжехвостые славки, белокрылый дятел и древесный гриб - пория пустынная. Этот тип биогеоценоза распространен в южной песчаной пустыне по долинным понижениям, характеризующимся повышенной гипсоносностью и уплотненностью песков, сравнительно неглубоким залеганием грунтовых вод (до 8-11 м) и большой контрастностью ритмических процессов в абиотической среде и биоте.

Рис. 6. Соотношение площадей парцелл в спелом черносаксаульнике с сомкнутыми кронами (а), в редкостойном илаковом (б) и в эфедровом (в, г, д) (южная песчаная пустыня, Восточные Каракумы)
Биогеоценосвязи выражаются в высокой степени закрепленности песков, образовании пятнистого фитогенного засоления их поверхностного слоя, локальных куполообразных поднятий грунтовых вод под деревьями черного саксаула, при этом стержнем и, вероятно, функционально активным элементом в подкроновых куполах увлажнения служит корень саксаула. Закрепление поверхности песков растительностью и повышенным засолением на подкроновых салфетках, а также некоторое известное торможение ветровых потоков довольно высоким и местами густым древостоем существенно ослабляет роль чисто эоловых процессов в формировании рельефа и в жизни биогеоценоза "черносаксауловых долин" Восточных Каракумов по сравнению с биогео-ценозами барханных и других типов полузакрепленных и слабо закрепленных песков (см. табл. 9). Вместе с тем зооэоловые процессы приобретают особенно большое значение в морфологии ландшафта и циклике развития ценоэлементарных структур. Немаловажное значение в этих процессах имеет роющая деятельность полуденной и большой песчанок и отчасти тонкопалого суслика.
| Природно-территориальные комплексы (фации) |
Количество кустарников на га, шт. |
Количество улавливаемого песка, кг |
| Черносаксаульник с сомкнутыми кронами | 400 | 0,001 |
| Поляна в черносаксаульнике (20X15 м) с молодыми всходами саксаула |
8 | 0,320 |
| Белосаксаульник с илаковым покровом | 143 | 0,430 |
| Белосаксаульник с нарушенным илаковым покровом | 340 | 3,600 |
| Белосаксаульник без илакового покрова (на гряде) | 364 | 10,730 |
| Переходная полоса от оголенных песков к белосаксаульникам без илакового покрова |
168 | 14,640 |
| Межбарханное понижение среди оголенных песков | 27 | 18,840 |
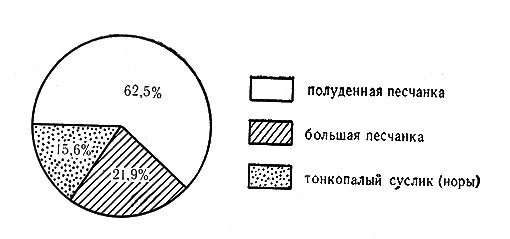
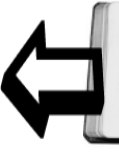
Рис. 7. Соотношение поселений разных видов грызунов-землероев в эфедровом черносаксаульнике (южная песчаная пустыня, Восточные Каракумы)
В целом физиономичность ландшафта долинных черносаксауловых лесов с эфедрой и морфология биогеоценоза этого типа определяются преимущественным развитием биотических процессов в отличие от биогеоценозов барханных и других слабо закрепленных песков, где, безусловно, преобладают абиотические процессы. Среди биогенных элементов ценоструктуры, которые можно обозначить как парцеллы (Дылис, Уткин, Успенская, 1963), важную функциональную роль играют подкроновые салфетки черного саксаула, сетевидная система солянково-злаковых бордюров подкроновых салфеток и зфедрогенные бугры (см. рис. 6, 7, 8).
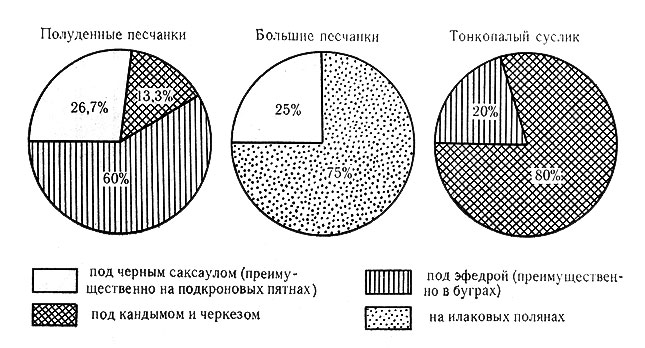
Рис. 8. Распределение поселений грызунов по парцеллам в эфедровом черносаксаульнике
Подкроновая салфетка черного саксаула - это функционально пескозакрепитель, стабилизатор солевого баланса, влагосохраняющее природное устройство, сезонно действующий термический рефугиум для наземных животных (беспозвоночных и позвоночных) и база развития "краевого эффекта" при контакте двух сред, на основе которого возникает сеть солянково-злаковых бордюров. Таким образом, подкроновые салфетки - это фактор создания множественности микроусловий в биогеоценозе и дифференциации среды жизни. Биопродукционным свойством подкроновая салфетка не обладает (см. рис. 9).
Рис. 9. Насыщенность парцелл эфедрового черносаксаульника норами полуденных песчанок и развитие ветроэрозионных процессов
Сетевидная система солянково-злаковых бордюров, возникающих по краю проекций крон, функционально - основной производитель полезно обращаемой части суммарной биопродукции биогеоценоза, утилизируемой животными, и вместе с тем травяные "бордюры" - место аккумуляции органического вещества, действующее перманентно, наконец, это участки развития максимальных контактов животных и растений и разных видов животных друг с другом (см. табл. 10). Таким образом, сетевидная система бордюров - зона повышенной активности биотических процессов в биогеоценозе, в том числе узел трофических цепей и база рассеяния и разноса органического вещества. Здесь концентрируются муравейники, жуки, полужесткокрылые и пауки (см. рис. 10).

Рис. 10. Почвенный профиль через подкроновое пространство черного саксаула
| Растительные сообщества | Надземная фитомасса, сухой вес кг/га | Численность | ||||||
| летние однолетники | злаки весенние | разнотравье | суммарный урожай весенне-летних трав | суммарная фитопродукция | позвоночных, особи/га | беспозвоночных, особи/100 м2 | ||
| птицы | большие песчанки | насекомые и пауки | ||||||
| Черносаксаульники илаково-эфедровые | 52 | 11 | 88 | 100-600 | 7860-54140 | 5,2 | 6-7* | 1000-2000 |
| Белосаксаульники илаковые | 10 | 60 | 23 | 110-430 | 4080 | 1,3 | 2-3 | 100-200 |
| Пионерная растительность барханных песков | - | - | - | - | 130-370 | 0,7 | 1 | 10-50 |
* (Численность среднего уровня. Высокая численность характеризуется в 2-3 раза большими показателями (Стальмакова, 1955).)
1 (По Р. К. Тогызаеву (1937), П. Д. Гунину (1973), Л. В. Комаровой (1974) и В. И. Кузнецову.)

Рис. 11. Почвенный профиль через эфедрогенный бугор и кривые суточных температур на поверхности (а) и на глубине 40 см (б)
Эфедрогенные бугры высотой от 0,2 до 2 м образованы опадом эфедры, сцементированным песком, чаще всего гипсоносным. Механический и химический состав песка в бугре дается в таблицах (табл. 11, 12). Бугры представляют собой плотные почвенные образования с большим числом разнообразных по размерам воздухоносных пор, которые обильно заселяются почвенными беспозвоночными, посещаются пауками и жуками (имаго). В буграх преимущественно поселяются полуденные песчанки. Здесь же происходит накопление и разложение биомассы погибших животных (см. рис. 11). Эфедрогенные бугры характеризуются свойством относительной термостабильности. В течение суток в летнее время колебания температуры внутри бугров наименьшие по сравнению со всеми другими точками биогеоценоза, зимой суточные колебания температуры и влажности по существу нивелируются.
| Горизонт, см | Соли, плотный остаток, % |
Размерный состав, мм | Физическая глина, % | Механический состав, по Качинскому |
||||
| 1-0,05 | 0,05-0,01 | 0,01-0,005 | 0,005-0,001 | <0,001 | ||||
| Под черным саксаулом (на подкроновом пятне) | ||||||||
| 0,5 | 0,27 | 82,12 | 11,86 | 0,76 | 2,33 | 2,93 | 6,02 | песчаный |
| 20 | 0,20 | 90,45 | 5,86 | 0,23 | 0,16 | 3,30 | 3,69 | песчаный |
| 40 | 0,20 | 93,48 | 1,70 | 0,53 | 0,43 | 3,86 | 4,82 | песчаный |
| 60 | 0,25 | 89,55 | 6,56 | 0,43 | 0,30 | 3,16 | 3,89 | песчаный |
| 80 | 0,19 | 93,95 | 2,83 | 0,16 | 1,66 | 1,40 | 3,82 | песчаный |
| 100 | 0,12 | 93,85 | 1,70 | 1,36 | 0,23 | 2,86 | 4,45 | песчаный |
| Под эфедрой (хорошо развитой) | ||||||||
| 0,5 | 0,15 | 77,71 | 16,6 | 0,76 | 2,13 | 2,30 | 5,69 | песчаный |
| 20 | 0,13 | 92,02 | 1,36 | 1,36 | 2,56 | 3,56 | 6,52 | песчаный |
| 40 | 0,08 | 94,55 | 1,73 | 0,93 | 0,56 | 2,23 | 3,72 | песчаный |
| 60 | 0,12 | 95,81 | 0,53 | 0,60 | 0,26 | 2,80 | 3,66 | песчаный |
| 80 | 0,09 | 95,04 | 1,66 | 0,50 | 0,10 | 2,70 | 3,30 | песчаный |
| 100 | 0,07 | 94,28 | 2,23 | 0,36 | 0,30 | 2,83 | 3,49 | песчаный |
| Под эфедрой (угнетенной) | ||||||||
| 0,5 | 0,12 | 75,65 | 19,93 | 0,16 | 0,96 | 3,30 | 4,42 | песчаный |
| 20 | 0,10 | 87,81 | 8,70 | 0,40 | 0,13 | 2,96 | 3,49 | песчаный |
| 40 | 0,19 | 89,41 | 7,63 | 1,43 | 0,10 | 1,43 | 2,96 | песчаный |
| 80 | 0,30 | 90,24 | 3,36 | 2,70 | 0,20 | 3,50 | 6,40 | песчаный |
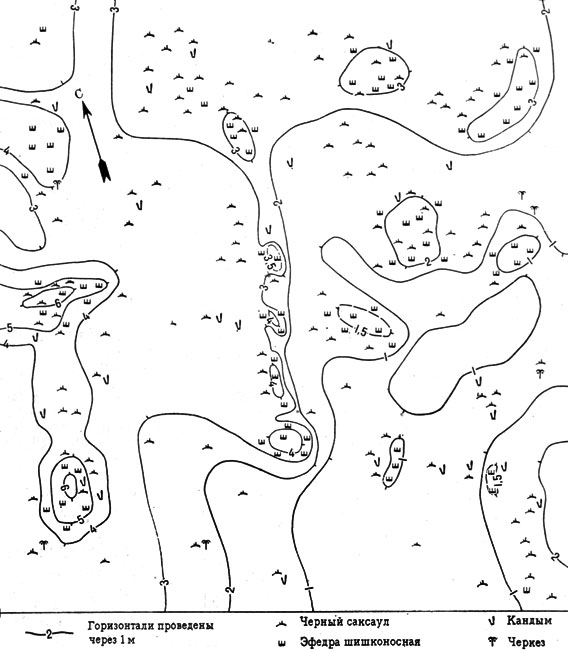
Рис. 12. Схема распределения древесно-кустарниковой растительности в эфедровом черносаксаульнике (песчаная пустыня южного типа, Восточные Каракумы. Пробная площадь 0,25 га)
В связи с этой особенностью эфедрогенные бугры, действующие как "подушки-термосы", функционально исполняют роль: 1) микробиотопа, максимально и перманентно наполненного почвенными животными (из числа которых складывается специфический животный комплекс эфедрогенных бугров); 2) места размножения некоторых наземных беспозвоночных и места обитания личиночных форм; 3) сезонного рефугиума для организмов, нуждающихся в периодических моментах термостабильности. Наряду с накоплением органического вещества животного происхождения в буграх происходит накопление, частичная консервация и переработка органических веществ растительного происхождения - в первую очередь опада эфедры и злакового разнотравья, развитого внутри куста эфедры по "крыше" и отчасти по скатам бугра и в значительной степени аналогичного покрову бордюров саксауловых подкроновых салфеток. Таким образом, от половины до 2/3 общей поверхности эфедрогенных бугров входит в систему сетевидных зон аккумуляции органического вещества биогеоценоза эфедрового черносаксаульника (см. рис. 12, 13).
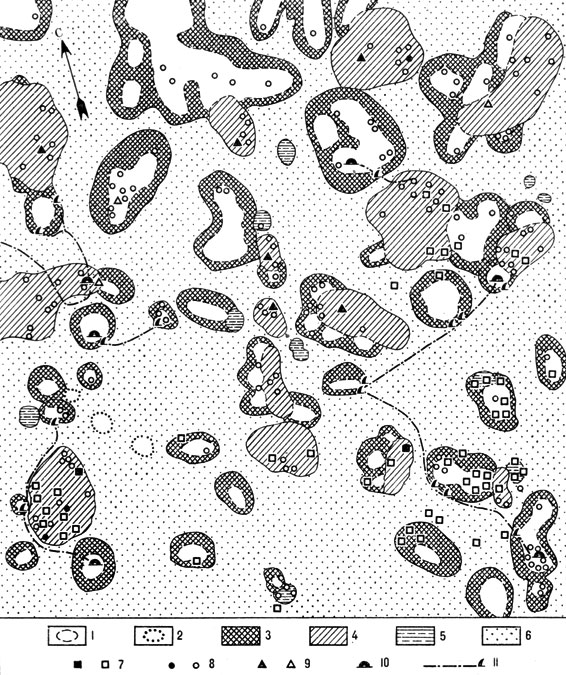
Рис. 13. Схема морфоструктуры биогеоценоза эфедрового черносак-саульника. 'Сетевидные зоны аккумуляции органического вещества', образованные подкроновой растительностью (песчаная пустыня южного типа, Восточные Каракумы. Пробная площадь 0,25 га): 1 - подкроновое пространство черного саксаула с повышенной засоленностью и уплотненностью поверхности почвы; 2 - 'реликтовое подкроновое пространство' на месте отмершего дерева; 3 - бордюрные микрогруппировки подкроновой растительности черного саксаула; 4 - подкроновые разнотравно-злаковые микрогруппировки эфедры шишконосной на эфедрогенных буграх; 5 - подкроновые солянково-злаковые микрогруппировки кандымов; 6 - песчаные поляны, заросшие илаком; 7 - выходы из нор (жилых и нежилых) большой песчанки; 8 - полуденной песчанки; 9 - тонкопалого суслика; 10 - муравейник; 11 - муравьиная тропа и место сбора колосков
| Горизонт, см |
Соли, плотный остаток, % |
Ионный состав | Сумма катионов |
|||||
| SO4" | HCO3' | Cl' | Ca¨ | Mg ¨ | Na·+K· | |||
| Под черным саксаулом (на подкроновом пятне) | ||||||||
| 0-5 | 0,27 | 1,33 | 1,78 | 0,37 | 0,17 | 1,39 | 1,92 | 3,48 |
| 5-20 | 0,20 | 0,67 | 1,36 | 0,62 | 3,35 | 0,35 | 1,98 | 2,65 |
| 20-40 | 0,20 | 0,50 | 1,36 | 0,75 | 0,26 | 0,53 | 1,83 | 2,65 |
| 40-60 | 0,25 | 1,64 | 0,96 | 1,12 | 0,43 | 0,17 | 3,12 | 3,72 |
| 60-80 | 0,19 | 0,72 | 0,92 | 0,75 | 0,35 | 0,52 | 1,52 | 2,39 |
| 80-100 | 0,12 | 0,48 | 0,58 | 0,62 | 0,35 | 0,35 | 0,97 | 1,68 |
| Под эфедрой (хорошо развитой) | ||||||||
| 0-5 | 0,15 | 0,29 | 0,22 | 0,50 | 0,87 | 0,17 | 0,57 | 1,01 |
| 5-20 | 0,13 | 1,04 | 0,44 | 0,25 | 0,43 | 0,35 | 0,95 | 1,73 |
| 20-40 | 0,08 | 0,29 | 0,54 | 0,37 | 0,35 | 0,26 | 0,59 | 1,20 |
| 40-60 | 0,12 | 0,98 | 0,49 | 0,37 | 0,26 | 0,43 | 1,14 | 1,84 |
| 60-80 | 0,09 | 0,38 | 0,45 | 0,50 | 0,61 | 0,35 | 0,37 | 1,33 |
| 80-100 | 0,07 | 0,102 | 0,52 | 0,50 | 0,52 | 0 | 0,50 | 1,12 |
| Под эфедрой (угнетенной) | ||||||||
| 0-5 | 0,12 | 0,89 | 0,40 | 0,37 | 0,70 | 3,35 | 0,61 | 1,66 |
| 5-20 | 0,10 | 0,58 | 0,49 | 0,37 | 0,43 | 0,43 | 0,58 | 1,44 |
| 20-40 | 0,19 | 0,79 | 0,34 | 1,25 | 1,65 | 0,26 | 0,47 | 2,38 |
| 40-80 | 0,13 | 0,27 | 0,37 | 1,50 | 1,13 | 0,68 | 0,33 | 2,14 |
Состав микрогруппировок подкроновой растительности эфедры включает в среднем около десятка видов злаков и разнотравья. В отличие от состава бордюрных микрогруппировок черного саксаула под эфедрой слабо развиваются однолетние солянки. Лондезия, как правило, не встречается, кохия Шренка - не типична. Микрогруппировки подкроновой растительности эфедры шишконосной (с распределением по экспозициям бугра) представлены ниже (на примере II фазы цикла).
Вершина бугра с "седловиной выдувания": Bromus tectorum, Salsola praecox, Senecio subdentatus, Malcolmia grandiflora, Cutandia memohitica, Carex physodes, Hyalea pulchella. Склон южной экспозиции: Bromus tectorum, Schismus arabicus, Senecio subdentatus, Delphinium camptocarpum, Salsola praecox, Malcolmia grandiflora, Hyalea pulchella, Carex physodes. Склон восточной экспозиции: Bromus tectorum, Delphinium camptocarpum, Hypecoum pendulum, Malcolmia grandiflora. Склон северной экспозиции: Bromus tectorum, Schismus arabicus, Silene папа, Senecio subdentatus, Delphinium camptocarpum, Salsola praecox, Carex physodes.
Кустарники на седловине выдувания (в зоне нор): Calligonum turkestanicum (1 особь, выc. 124 см), С. саput medusae (1 особь, выc. 166 см). Склон северной экспозиции (в зоне нор): С. caput medusae (1 особь, выc. 162 см). Склон восточной экспозиции (в периферийной зоне нор): Salsola richteri (1 особь, выc. 132 см). Особенности состава микрогруппировок зависят от экспозиции склонов бугра и нарушений поверхности почвы роющей деятельностью полуденных песчанок. Последние содействуют проникновению на эфедрогенный бугор более псаммофильных видов травянистых растений, в том числе песчаной осоки (Carex physodes) и кустарников. Так, в местах пескоройной деятельности песчанок, в "седловинах выдувания" на буграх эфедры шишконосной поселяются кандымы: туркестанский, щетинистый, голова Медузы, солянка Рихтера (черкез) и подрост черного и белого саксаулов.
Что касается топографии микрогруппировок растений и парцелл подкронового пространства черного саксаула, то следует отметить их концентрическое расположение в отношении ствола дерева и собственную насыщенность каждой из парцелл специфическими группировками организмов, связанных именно со своей парцеллой. Как уже отмечалось, муравьи и некоторые виды пауков концентрируются только в зоне "бордюрной растительности" по краю проекции кроны, а личинки саксаулового усача и гриб (Poria deserlorum) - преимущественно в зоне корневой шейки ствола и в верхней части корня дерева и т. п.
Такое размещение организмов, когда отдельные структурные элементы биогеокомплекса имеют еще и свою отчасти автономную структуру, подтверждаемую устойчивыми ценосвязями, можно назвать "системой вложенных структур". Подобный тип организации биогеоценозов характерен для южных пустынь с их высокой аридностью среды, что определяется возрастанием биологической ценности условий защищенности организмов, получающих лучшее развитие под покровом растений, изменяющих вокруг себя микроусловия ("покровный эффект").
| Виды | Количество особей, шт. | Виды | Количество особей, шт. | Виды | Количество особей, шт. |
| Учет 13 октября |
* | Учет 14 октября |
* | Учет 15 октября |
* |
| Такыры в долине р. Сумбар | |||||
| Zabrus morio Mén | 2 | Zabrus morio Mén | 1 | Chrysomelidae | 1 |
| Amara (Amathitis) dissors Tschitsch | 1 | Cicindellidae | 1 | Chactocnema sp. | 3 |
| Opatroides punctulatus Brulle | 2 | C. aenescens | 7 | Camponotus fedtshenkoi | 1 |
| Dissonomus tibialis Rtt. | 2 | Messor sp. | 6 | Messor sp. | 1 |
| Bothynoderes strabus | 1 | Monomorium sp. | 1 | Engistus exsangius | 5 |
| Macrotarrhus sp. | 2 | Simuliidae | 1 | Anthemina pusio Kol. | 1 |
| Cataglyphis jomud K. Arn. | 1 | Lepidoptera | 1 | Miridae sp. | 1 |
| Messor sp. | 2 | Engistus exsangius Stal. | 1 | Thysanura sp. | 1 |
| Empusa pennicornis (lavra) | 1 | Cicadelidae | 1 | Araneina | 3 |
| Stictopleurus angustus Rent. | 1 | Anacanthotermes sp. | 3 | * | * |
| Araneina | 1 | Araneina | 3 | * | * |
| Hemilepistus | 3 | Hemilepistus | 7 | * | * |
| Всего | 19 | * | 34 | * | 17 |
| Такыры приморской пустыни | |||||
| Adesmia karelini Fisch. w. | 1 | Metabletus parallelus Rt. | 1 | Amara (Harpalodema) ruthena Tschitsch | 2 |
| Opatroides punctulatus Brulle. | 1 | Bothynoderes sp. | 2 | A. (Harpalodema) ahngeriana Tschitsch | 1 |
| Dissonomus tibialis Rtt. | 1 | Camponotus turcestanicus | 4 | Aphodius sp. | 1 |
| Camponotus turcestanicus | 6 | Monomorium sp. | 1 | Bradynema signata Jak. | 3 |
| Cataglyphis setipes | 1 | Scantius aegyptius L. | 1 | Hyalocoris pilicornis Jak. | 1 |
| Monomorium sp. | 1 | Cercinthus lehmani Kol. | 1 | Engistus exsangius Stal. | 3 |
| Centrocoris voixemi Put. | 3 | Horventhiolus syriacus Rent. | 2 | Cymophyes golodnajana Stal. (на злаке Aeluropus litoralis) | 2 |
| Emblethis dilaticolis Jak. | 1 | Gryllidae | 2 | Centrocoris volxemi Put. | 1 |
| Engistus exsangius Stal. | 2 | Olpium sp | 1 | Araneina | 1 |
| Gryllidae | 1 | * | * | * | * |
| Epitetranichus sp. | 2 | * | * | * | * |
| Hemilepistus sp. | 6 | * | * | * | * |
| Scutigera sp. | 1 | * | * | * | * |
| Araneina | 2 | * | * | * | * |
| Всего | 27 | * | 15 | * | 17 |
1 (Определение беспозвоночных проведено в Зоологическом институте АН СССР О. Л. Крыжановским, Г. С. Медведевым и И. М. Кержнером.)
Биогеоценоз черносаксаульников Каракумов организован по типу "системы вложенных структур". То же относится к группировкам растений и животных, развивающихся под покровом кустов крупных солянок (группы Salsola crassa), на солончаках и такыровидных участках в речных долинах. Например, в долине р. Сум-бар наблюдались группировки беспозвоночных, возникшие под защитой солянок и включавшие комплексы видов, живущих на самой солянке, на маленьких куртинках злака ажрека (Aeluropus litoralis Pall.), растущего вокруг выбросов из норок землероющих мокриц рода Неmilepistus под кроной куста солянки, и животных, населяющих норки мокриц, где сохраняется влажность и где, следовательно, могут обитать даже мезофильные организмы. В табл. 13 представлен перечень видов беспозвоночных животных, существующих под покровом солянок на такырах в Западной Туркмении.