

Подкласс растительные жгутиконосцы, или фитомастигины (Phytomastigina)
Многим, вероятно, приходилось видеть в природе своеобразное явление "цветения воды", когда вода в луже, пруду или озере становится изумрудно-зеленой. Причиной "цветения" являются различные микроскопические растительные организмы, развивающиеся в несметных количествах. Очень часто это бывают разные представители фитомастипш.
Хламидомонады. Эти одноклеточные организмы часто являются причиной "цветения". К роду хламидомонада (Сhlamydomonas) относится свыше 150 видов. Тело большинства их яйцевидное и шаровидное (рис. 41), очень небольших размеров (10-30 мк). Все тело покрыто довольно прочной оболочкой из целлюлозы - вещества, из которого состоят оболочки растительных клеток. На переднем конце расположены два жгутика одинаковой длины. Ярко-зеленый цвет жгутиконосца обусловлен наличием в цитоплазме его чашевидного хроматофора, несущего зеленый пигмент -хлорофилл. Весь хлорофилл сосредоточен в хроматофоре и вне его никогда не встречается. В области прикрепления жгутиков имеется одна или две сократительные вакуоли - это органоид осморегуляции и выделения. У некоторых видов хламидомонад на переднем конце тела имеется еще одно образование в виде небольшого пятна яркого красно-коричневого цвета. Этостигма, илиглазок. Его окраска обусловлена наличием особого пигмента, называемогогематохромом. Хламидомонады обладают резко выраженным положительным фототропизмом: они всегда плывут в сторону источника света. Некоторые наблюдения позволяют считать, что стигма к представляет собой органоид, связанный с восприятием светового раздражения, и является по своей функции действительно клеточным глазком. В цитоплазме обычно присутствуют зерна крахмала, являющиеся продуктом фотосинтеза.
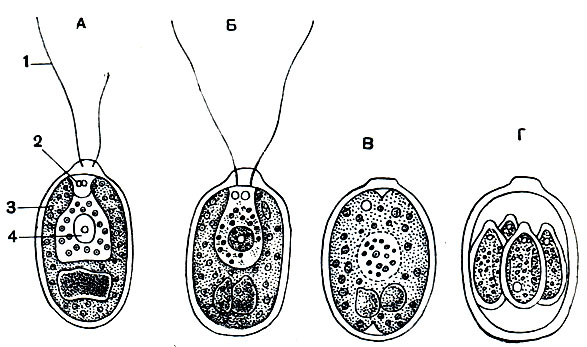
Рис. 41. Хламидомонада Chlamydomonas angulosa: А - неделящаяся особь; Б - Г - стадии деления. 1 - жгутик; 2 - сократительная вакуоля; з - хромато-фор; 4 -ядро
Клеточное ядро хламидомонад сферической формы и расположено примерно в середине клетки.
Размножение большинства хламидомонад происходит в неподвижном состоянии (рис. 41): жгутиконосец теряет жгутики, опускается на дно. Внутри оболочки происходит деление протоплазматической части клетки (протопласта) сначала на две, затем на четыре части. Через разрыв оболочки материнской клетки образовавшиеся в результате деления дочерние клетки выходят наружу, приобретают жгутики, дорастают до величины материнской клетки и выделяют оболочку. Рассмотренный только что процесс представляет собойбесполое размножение.
Кроме бесполого размножения, у большинства хламидомонад известенполовой процесс. Он сводится к тому, что вегетативные клетки становятся гаметами, которые попарно сливаются. Слияние гамет сопровождается слиянием ядер. Образовавшаяся в результате оплодотворениязигота теряет жгутики и покрывается прочной оболочкой. Внутри нее происходит деление протопласта на четыре клетки, у которых развиваются жгутики. Клетки превращаются в четыре особи хламидомонад, переходящих к свободноподвижному образу жизни.
Колониальные фитомастигины. Среди фитомастигин есть немало колониальных видов. Колонии образуются в результате бесполого размножения путем деления, при котором продукты деления остаются связанными друг с другом. Таким образом возникает как бы особь высшего порядка, включающая в себя некоторое (иногда очень значительное) число отдельных клеток, каждая из которых соответствует одной хламидомонаде. Рассмотрим некоторые наиболее широко распространенные виды колониальных фитомастигин.
В пресных водах часто встречаются маленькие изящные колонии гониумов (Gonium pectorale), представляющие собой зеленые пластиночки, каждая из которых состоит из 16 клеток (рис. 42). Клетки колонии по своему строению полностью соответствуют хламидомонадам. Отдельные клетки связаны друг с другом бесцветной прозрачной студенистой массой. Эта масса образуется в результате ослизнения наружного слоя оболочек клеток. Благодаря биению жгутиков вся колония активно плавает в воде.
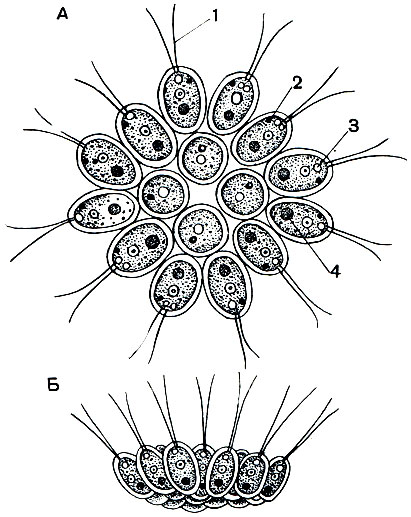
Рис. 42. Колониальная фитомастигина Gonium pectorale: 1 - жгутики; 2 - глазок (стигма); 3 - сократительная вакуоля; 4 - ядро
Другой часто встречающейся пресноводной колониальной формой фитомастигин является эвдорина (Eudorina elegans). Колонии этого вида, в отличие от гониумов, шаровидны (рис. 43) и в большинстве случаев слагаются из 32 клеток (изредка число клеток колонии может быть и меньше, а именно 16 и 8). Как и у гониумов, отдельные особи колонии эвдорин связаны между собой студенистым прозрачным веществом. Диаметр колоний этого вида варьирует в довольно широких пределах (от 50 до 200 мк).
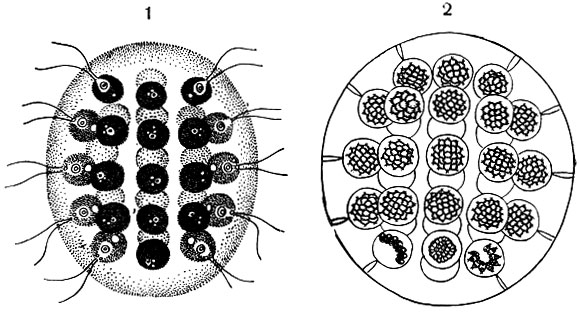
Рис. 43. Шаровидная колония Eudorina elegans: 1 - колония из 32 клеток; 2 - бесполое размножение (каждая особь колонии в результате последовательных делений дает начало новой, дочерней колонии)
Каждая клетка колоний гониумов и эвдорин сохраняет способность к бесполому размножению, которое удается наблюдать довольно часто. К делению приступают одновременно все клетки колонии (рис. 43, 2). Каждая клетка у гониума претерпевает 3, а у эвдорин - 4 последовательных, быстро следующих друг за другом деления. В результате внутри общей слизистой оболочки колонии образуются новые, дочерние колонии, число которых соответствует числу приступивших к размножению клеток. В дальнейшем слизистый чехол материнской колонии распадается, и дочерние колонии приступают к самостоятельному существованию.
Очень большой сложности достигает строение колоний у вольвокса (Volvox). Несколько видов этого рода - частые обитатели небольших пресноводных водоемов (рис. 44).
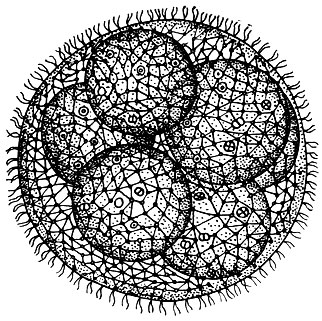
Рис. 44. Колония Volvox aureus с дочерними колониями внутри материнской
Вольвоксы образуют крупные шаровидные колонии. У наиболее часто встречающегося вида Volvox aureus диаметр колоний равен 500-850 мк, а размеры колонии Volvox globator достигают 2 мм. У Volvox aureus в состав колонии входит 500-1000 отдельных клеток (рис. 44), а у Volvox globator - до 20 тыс. Основная масса колонии состоит из студенистого веществаг образующегося в результате ослизнения клеточных оболочек. Самый наружный слой его представляет собой особый плотный кожистый слой, придающий всей колонии значительную прочность. Центральные части студенистого вещества гораздо менее плотной консистенции - они полужидкие. Отдельные клетки колонии располагаются в самом периферическом слое. Каждая клетка имеет в основных чертах такое же строение, как и одиночные хламидомонады. В колонии вольвокса отдельные особи не вполне изолированы друг от друга, они связаны между собой тончайшими цитоплазматическими мостиками (рис. 45).
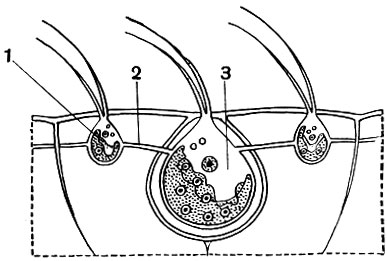
Рис. 45. Небольшой участок колонии Volvox aureus (схематизированно): 1 - вегетативная особь колонии; 2 - цитоплазматический мостик; 3 - более крупная особь, в результате развития которой получаются новые, дочерние колонии
Движение вольвокса осуществляется благодаря совместному действию жгутиков. Несмотря на то что колония шаровидна, она обладает некоторой полярной диффе-ренцировкой. Последняя выражается в том, что на одном полюсе стигмы (глазки) развиты сильнее, чем на другом. Движение вольвокса тоже оказывается ориентированным: он плывет вперед тем полюсом, на котором глазки сильнее развиты. Мы видели, что у относительно просто устроенных колоний, таких, как гониум и эвдорина, все клетки способны путем бесполого размножения давать начало новой колонии. У вольвокса дело обстоит иначе. Огромное большинство клеток колонии не способно делиться. Этим свойством обладают лишь очень немногие особи. Число их в колонии относительно невелико: чаще всего 4-10. Они располагаются в большинстве своем в нижней части колонии (в той части ее, где глазки относительно слабо развиты). Эти клетки, называемые "вегетативными клетками размножения", удается рассмотреть лишь в относительно молодых колониях. В более старых они находятся на различных стадиях развития в дочерние колонии. Не вдаваясь в детали этого довольно сложного процесса, укажем, что дочерние колонии развиваются внутри материнской в результате многократного деления "вегетативных клеток размножения" (рис. 46). Когда размеры дочерних колоний увеличатся настолько, что они уже не смогут поместиться внутри старой, последняя лопается и погибает, а дочерние колонии выходят наружу. Летом, в период энергичного размножения, нередко можно наблюдать, как внутри дочерних колоний, находящихся еще внутри материнской, начинают развиваться внучатые колонии. Кроме бесполого размножения, у колониальных фитомастигин наблюдается и половой процесс, который протекает у различных видов по-разному. Укажем лишь, что при половом процессе в колонии образуются гаметы, в результате слияния (оплодотворения) которых получается зигота. Из зиготы путем ряда последовательных делений развивается новое поколение колоний.
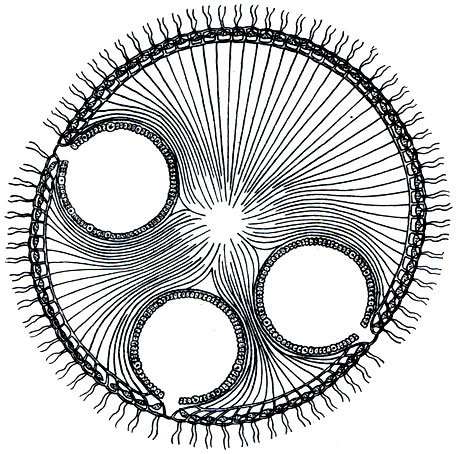
Рис. 46. Разрез через колонию вольвокса с тремя дочерними колониями внутри материнской (схематизированно)
Отряд панцирные жгутиконосцы, или перидинеи (Dinoflagellata, или Peridinea)
Панцирные жгутиконосцы - обширная группа простейших, широко распространенная в морях и пресных водах. Подавляющее большинство видов этого отряда относится к планктонным формам.
Характерной особенностью панцирных жгутиконосцев является своеобразное строениежгутикового аппарата (рис. 47). Имеющиеся у них два жгута берут начало рядом друг с другом на одной стороне тела простейшего, которую условно называют брюшной. Один жгутик направлен назад и свободно выдается в окружающую среду. Начальная часть его лежит в небольшом желобке, идущем вдоль тела. Второй жгутик, называемый поперечным, опоясывает все тело в экваториальной плоскости и расположен в довольно глубоком желобе, называемом поясом (cingnlum), который опоясывает все тело. Поперечный жгут производит характерные волнообразные движения, что создает ложное впечатление, будто в поперечной борозде расположен ряд ресничек.
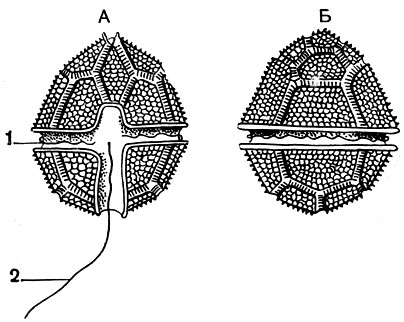
Рис. 47. Панцирный жгутиконосец Peredinium tabulatum: А - вид с брюшной стороны; Б - вид со спинной стороны. 1 - поперечный жгутик в пояске; 2 - свободный жгутик
Вторая характерная особенность большинства (но не всех) панцирных жгутиконосцев - наличие у нихоболочки, состоящей изклетчатки. Эта оболочка складывается из строго определенного числа закономерно расположенных отдельных пластинок, соединенных друг с другом швами.
Цитоплазма жгутиконосцев обычно бывает окрашена в буроватый цвет, который обусловлен наличием в ней большого количества мелких дисковидных хроматофоров, содержащих хлорофилл и некоторые дополнительные пигменты. Продуктом ассимиляции является крахмал, зерна которого рассеяны в цитоплазме.
Рассмотрим представителей двух наиболее широко распространенных родов динофлагеллят.
На рисунке изображен часто встречающийся в пресных водах перидиний (Peridinium tabulatum). У него отчетливо выражены упомянутые выше пластинки. Форма тела более или менее правильно яйцевидная. Иногда бурное развитие перидиний вызывает "цветение" воды.
Широко распространены как в пресной, так и в морской воде многочисленные виды рода церациум (Ceratium). Характерной особенностью представителей этого рода являются длинные выросты. У пресноводного Ceratium hirudinella (рис. 48) один вырост (апикальный) направлен вперед и три (антапикальных) - назад.
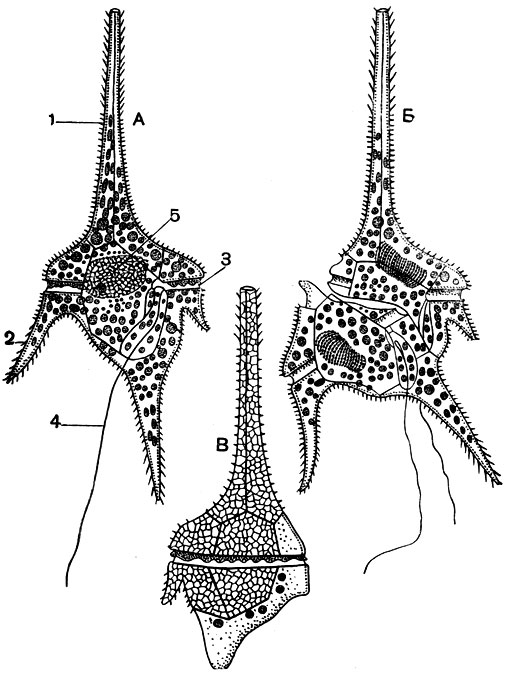
Рис. 48. Ceratium hirudinella: А - неделящаяся особь; Б - деление; В - только что разделившаяся особь, часть пластинок на заднем конце недоразвита. 1 - апикальный вырост; 2 - антапикальный вырост; 3 - поясок; 4 - задний жгутик; 5 - ядро
Особенно сильно эти выросты выражены у морских видов Ceratium, у которых они вторично ветвятся (рис. 49). Для морских же видов характерно образование временных колоний-цепочек (рис. 50), которые возникают в результате того, что разделившиеся особи остаются некоторое время связанными друг с другом. Длинные отростки у морских видов Ceratium представляют собой приспособление к планктонному образу жизни. Отростки увеличивают поверхность тела, что способствует "парению" в воде. Аналогичные приспособления мы уже видели у радиолярий.
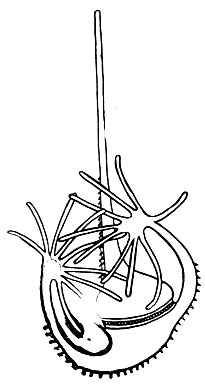
Рис. 49. Морской панцирный жгутиконосец Ceratium palmatum
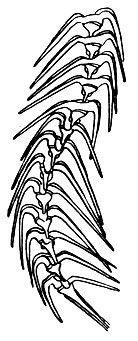
Рис. 50. Морской панцирный жгутиконосец Geratium voltur. Цепочка, образовавшаяся в результате делений
Особенно причудливую форму приобретает панцирь у представителей рода Ornithocercus, где по краям борозд образуются длинные крыловидные выросты (рис. 51). Это тоже приспособление к "парению" в воде.
В планктоне теплых и тропических морей очень часто, и нередко в огромных количествах, встречается жгутиконосец ночесветка (Noctiluca miliaris). В летнее время это простейшее обычно развивается и в Черном море.
Ночесветка имеет шаровидное тело до 2 мм в диаметре. В отличие от других панцирных жгутиконосцев, она не имеет оболочки из клетчатки и лишена хроматофоров (рис. 52). Поэтому ночесветка не способна к фотосинтезу. В отличие от большинства Dinoflagellata, ночесветке свойственно животное (анимальное) питание. Она заглатывает и переваривает различные оформленные пищевые частицы: мелкие водоросли, мелкие виды простейших.
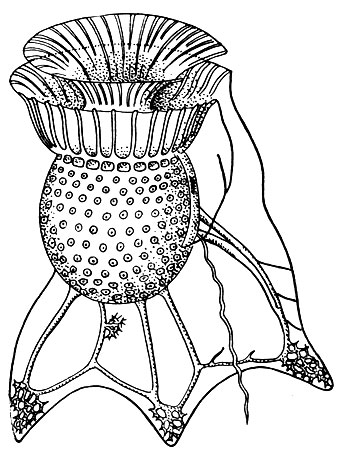
Рис. 51. Панцирный жгутиконосец Ornithocercus magnificus
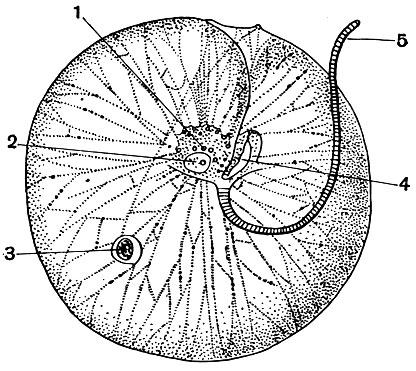
Рис. 52. Ночесветка Noctiluca miliaris: 1 - центральное скопление цитоплазмы; 2 - ядро; 3 - пищеварительные вакуоли; 4 - жгутик, расположенный в области ротового углубления; 5 - жгутик 'щупальце'
На одной стороне шаровидного тела ночесветки имеется углубление, соответствующее продольному желобку типичных панцирных жгутиконосцев. В глубине его помещается отверстие, выполняющее функцию рта. Через него и происходит поглощение пищи. В области этого же углубления помещаются два жгута. Один из них короткий и толстый. Он скорее напоминает щупальце. Этот жгут совершает относительно очень медленные колебательные движения (не более 10 в минуту), способствующие направлению пищевых частиц к ротовому отверстию. Второй, более короткий и тонкий жгутик также расположен в области ротового углубления (см. рис. 52, 4).
Очень своеобразно строение эндоплазмы ночесветки. Непосредственно под ротовым отверстием располагается центральное скопление цитоплазмы, от которого к периферии во всех направлениях отходят тонкие тяжи. Вся же остальная масса тела заполнена жидкостью и представляет собой систему вакуолей. Ядро расположено в центральном скоплении цитоплазмы. В цитоплазматических тяжах имеются многочисленные жировые включения.
Описанное выше своеобразное строение цитоплазмы следует рассматривать как одну из форм приспособления к планктонному образу жизни. Сильная вакуолизация цитоплазмы и наличие жировых включений ведут к уменьшению удельного веса и, следовательно, способствуют "парению" в толще воды.
У ночесветки есть еще одна интересная особенность, за которую она и получила свое название. При раздражении (механическом, химическом и т. п.) ночесветка ярко "вспыхивает". Это явление незаметно днем, при ярком свете, но очень отчетливо выражено в темноте, ночью. Ночесветка - это один из организмов, вызывающих явление свечения, столь характерное для теплых морей. Всякий, кому приходилось в теплую летнюю ночь плыть по Черному морю в лодке или на пароходе, вероятно, имел возможность наблюдать это эффектное явление. При ударе весел по воде, при падении капель воды с весел в море, при вращении пароходного винта и т. п. вода начинает светиться слабым фосфорическим, но вполне отчетливым светом. При этом видны бывают отдельные вспышки, напоминающие искорки.
Свечение наступает в результате окисления жировых включений, о наличии которых уже говорилось выше.
Свечение имеет, вероятно, защитное значение (отпугивает хищников).
Некоторые виды панцирных жгутиконосцев являются симбионтами радиолярий. Недавно выяснено, что они в большом количестве поселяются и в мягких тканях коралловых полипов.
Отряд эвгленовые (Euglenoidea)
Значительный интерес представляет отряд эвгленовых (Euglenoidea), так как в пределах этой группы встречаются все типы питания - от типично растительного (аутотрофного) до типично животного (анимального). Многочисленные виды рода эвглена (Euglena) широко распространены в пресноводных водоемах, особенно часто они встречаются в загрязненных водах. Строение эвглен представлено на рисунках 40 и 53. Эвглены снабжены одним жгутиком. В передней части тела их расположена сократительная вакуоля и ярко-красный глазок (стигма). В цитоплазме содержатся зеленыехроматофоры, несущиехлорофилл. Форма хроматофоров у разных видов различна. Наружный слой цитоплазмы образуетпелликулу. У многих видов она бывает структурирована: несет расположенные рядами утолщения (рис. 53). Тело некоторых эвглен покрыто очень тонкой эластичной пелликулой. Эти организмы способны сокращаться и вытягиваться по продольной оси. У эвгленовых имеется одно ядро с крупным ядрышком.
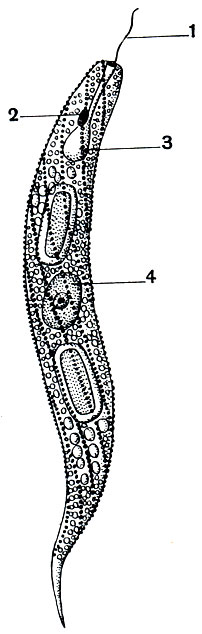
Рис. 53. Euglena spirogyra. Пелликула структуирована. 1 - жгутик; 2 - глазок (стигма); 3 - резервуар сократительной вакуоли; 4 - ядро
Замечательной особенностью многих видов эвглен является способность менять характер питания и обмена веществ в зависимости от условий среды. На свету при наличии в окружающей среде минеральных солей, содержащих необходимые химические элементы, эвгленам свойствен типичный аутотрофный обмен. В их теле протекает фотосинтез и усвоение неорганических солей. Если тех же эвглен поместить в темноту, в раствор, содержащий орнические вещества. Таким образом эвглены переходят от аутотрофного к гетеротрофному питанию. Гетеротрофное питание осуществляется у них не путем заглатывания оформленных частиц пищи, а путем осмоса: поглощения растворенных в окружающей среде органических питательных веществ через пелликулу. Такой способ питания называютсапрофитным. Часто, развиваясь в загрязненных водах, где имеется большое количество растворенных органических веществ, эвглены сочетают оба типа питания - аутотрофный и сапрофитный. Если обесцвеченных, культивируемых в темноте эвглен вновь перенести на свет, то через короткий промежуток времени они вновь зеленеют и переходят к аутотрофному питанию. Таким образом, мы видим, что на низших ступенях развития органического мира животный и растительный тип обмена нерезко отграничены друг от друга.
В отряде эвгленовых имеются роды и виды, которые окончательно утеряли способность к аутотрофному типу обмена веществ и при всех условиях не синтезируют хлорофилл. Некоторые из них, способные заглатывать оформленные частицы пищи, перешли уже к настоящему животному типу питания и обмена веществ (например, роды Peranema, Urceolus).
Подкласс животные жгутиконосцы, или зоомастигины (Zoomastigina)
Среди зоомастигин имеется значительное количество видов, живущих в морской и пресной воде и питающихся либо сапрофитно (осмотическим путем), либо путем заглатывания оформленных частиц пищи (анимальное питание). Однако большая часть зоомастигин перешла к паразитическому образу жизни в различных растительных и животных организмах. Рассмотрим сначала некоторых наиболее характерных и широко распространенных представителей свободноживущих зоомастигин.
В сильно загрязненных органическими веществами водах почти всегда в больших количествах можно встретить разные виды рода Bodo, относящегося к отряду протомонад (Protomonadina). Это очень мелкие жгутиконосцы (рис. 54) длиной 10-25 мк, с двумя жгутиками. Один из них направлен вперед и активно движется, вызывая поступательное движение простейшего. Второй жгутик направлен назад и более или менее пассивно волочится в воде. Он выполняет, вероятно, функцию руля. Форма тела яйцевидная, довольно сильно варьирующая. Ядро одно, расположено в передней трети тела. У основания жгутиков имеется довольно крупное сферической формы образование, называемое парабазальным телом. Предполагают, что в нем сосредоточены вещества, потребляемые жгутиком в процессе его движения.
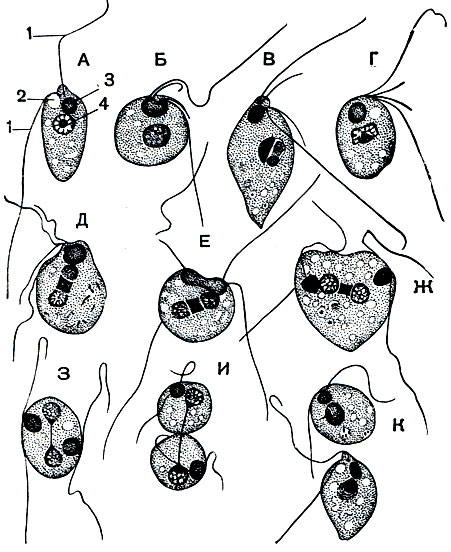
Рис. 54. Bodo edax. А - неделящаяся особь; Б - К - последовательные стадии деления. 1 - жгутики; 2 - сократительная вакуоля; 3 - парабазальное тело; 4 - ядро
Bodo свойствен анимальный способ питания. У основания жгутиков расположен небольшой участок цитоплазмы, лишенный пелликулы. В этой области и происходит заглатывание пищи, каковой обычно являются бактерии. В передней трети тела (на уровне парабазального тела) расположена сократительная вакуоля.
Большой интерес для понимания связей между саркодовыми и жгутиконосцами представляют простейшие, объединяемые в группу ризомастигин (Rhizomastigina), которая некоторыми зоологами рассматривается как самостоятельныйотряд илиподотряд жгутиконосцев, а другими (например, известным французским зоологом Грассе) включаетсяв класс саркодовых. Это расхождение во мнениях не случайно. Оно объясняется тем, что ризомастигины сочетают в своем строении признаки обоих классов: у них имеются и псевдоподии и жгутик (рис. 55). На прилагаемом рисунке изображено строение одного из представителей этой группы - Mastigamoeba aspersa. На переднем конце тела расположен жгутик, а на поверхности всего тела - хорошо развитые многочисленные псевдоподии, несущие, особенно на заднем конце, щетинкоподобные выросты. Вся цитоплазма отчетливо разделена на два слоя: наружную (светлую) эктоплазму и внутреннюю (зернистую) эндоплазму. Ядро помещается на переднем конце тела, сократительная вакуоля - на заднем. Размеры Mastigamoeba aspersa достигают 100 мк. Встречаются эти организмы не очень часто, преимущественно в болотных, богатых гумусовыми веществами водах.
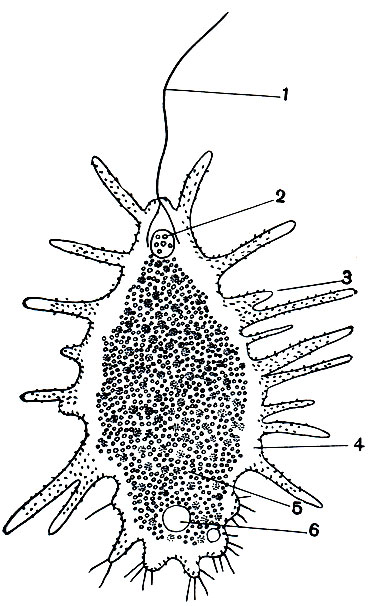
Рис. 55. Жгутиконосец Mastigamoeba aspersa, обладающий жгутиком и псевдоподиями: 1 - жгутик; 2 - ядро; 3 - псевдоподии; 4 - эктоплазма; 5 - эндоплазма; б - сократительная вакуоля
Наличие таких форм, как различные ризомастигины, показывает, что между саркодовыми и жгутиконосцами существуют тесные связи, что эти два класса связаны филогенетически.
Паразитические жгутиконосцы, как уже говорилось выше, многочисленны и разнообразны. Мы остановимся лишь на немногих представителях, наиболее интересных в практическом или биологическом отношении. Рассматривать мы их будем не в порядке зоологической системы, а по месту и характеру паразитирования.
Жгутиконосцы, паразитирующие в кишечном тракте человека и позвоночных животных. Кишечник позвоночных, а также и человека является местом обитания довольно многих видов жгутиконосцев. Рассмотрим некоторых из них. Широко распространен род трихомонад (Trichomonas), обладающий многочисленными видами. В кишечнике человека паразитирует Trichomonas hominis (рис. 56). Местом локализации являются тонкие и толстые кишки. Строение его, несмотря на небольшие размеры (7 -10 мк в длину, 4-5 мк в ширину), довольно сложно. Тело почти округлое. На переднем конце расположены 4 жгутика, каждый из которых берет начало от базального зерна (блефаропласта). Пятый жгутик заворачивает назад и идет вдоль края тела, прирастая к нему при помощи тонкой цитоплазматической перепонки. Лишь самый задний конец его торчит свободно. Этот пятый жгутик вместе с перепонкой образует так называемуюундулирующую мембрану, которая находится в постоянном волнообразном движении и играет существенную роль в движении самого жгутиконосца. Через всю цитоплазму от области расположения базальных зерен до самого заднего конца проходитаксостиль. Он представляет собой опорную эластичную нить, которая является внутренним скелетом простейшего. Наличие его свойственно и многим другим паразитическим жгутиконосцам. Трихомонады живут в содержимом кишечника, обладающем довольно густой консистенцией, им приходится передвигаться в этой среде. Одно ядро помещается на переднем конце тела.
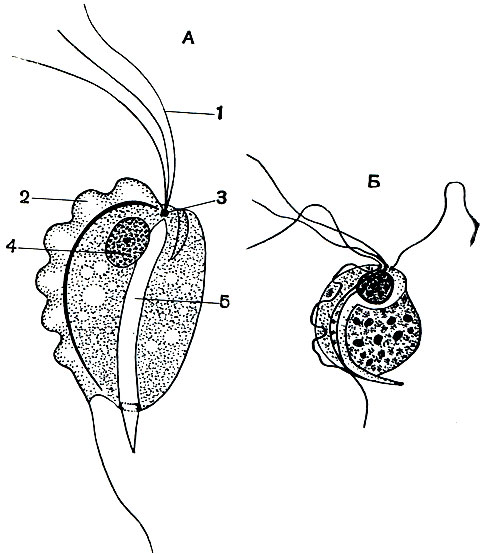
Рис. 56. Жгутиконосцы рода Trichomonas: А - схема строения; Б - Trichomonas hominis из кишечника человека. 1 - жгутики; 2 - ундулирующая мембрана; 3 - базальные зерна жгутиков; 5 - аксостиль
У трихомонад имеется ротовое отверстие, расположенное на переднем конце, вблизи основания жгутиков, на стороне, противоположной той, по которой проходит ундулирующая мембрана. Размножаются трихомонады продольным делением.
В кишечнике человека трихомонады встречаются довольно редко. При массовом размножении могут, по-видимому, вызывать понос.
Трихомонады встречаются, кроме кишечника человека, в кишечнике многих позвоночных животных, в частности кроликов, мышей, ящериц.
В верхних отделах тонкого кишечника человека довольно часто встречается еще один вид паразитических жгутиконосцев -лямблии (Lamblia intestinalis, рис. 57). Этот вид обладает весьма сложным строением. Форма тела грушевидная, заостряющаяся к заднему концу. Длина около 15 мк, ширина в передней части 7-8 мм. Все строение лямблии строго двустороннесимметрично. По продольной длинной оси ее можно рассечь на две зеркально равные половинки. Все тело довольно сильно сплющено в спинно-брюшном направлении. Спинная сторона выпуклая, брюшная плоская. В передней части брюшной поверхности лямблий имеется углубление, представляющее собой присоску, при помощи которой жгутиконосец плотно присасывается к эпителиальным клеткам кишечника (рис. 57). Посередине тела до самого заднего конца проходит двойной аксостиль, состоящий из двух эластичных нитей. Имеется четыре пары жгутиков, расположение которых ясно видно на рисунках. Задняя пара их является продолжением аксостилей. Ядер два, они лежат справа и слева от аксостилей в области присоски.
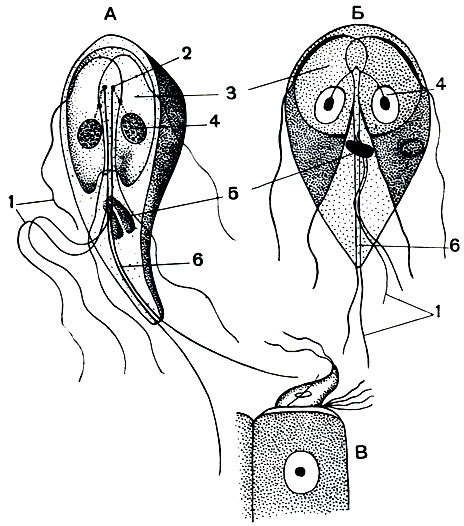
Рис. 57. Паразитирующий в кишечнике человека жгутиконосец Lamblia intestinalis: А - вид сбоку; Б - вид с брюшной стороны; В - лямблия, присосавшаяся к эпителиальной клетке. 1 - жгутики; 2 - базальные зерна; 3 - присоска; 4 - ядро; 5 - парабазальное тело; 6 - аксостиль
Лямблии - очень нежелательные "гости" кишечника человека. Нередко заражение ими проходит без каких-либо болезненных симптомов. Вместе с тем известны многочисленные случаи, когда заражение лямблиями сопровождалось кишечными расстройствами. Через желчный проток из двенадцатиперстной кишки лямблии могут проникать в желчный пузырь, вызывая заболевания типахолецистита. У человека лямблии гораздо чаще встречаются у детей, чем у взрослых.
Попадая в нижние отделы кишечника, лямблии могут инцистироваться: они теряют жгутики и одеваются оболочкой. В состоянии цист они и выходят из кишечника наружу. Человек заражается, проглатывая цисты лямблий.
Лямблии встречаются и у многих млекопитающих, в частности у кроликов и мышей. Однако у каждого вида хозяина существует свой, свойственный только ему вид лямблий. Лямблии кролика или мыши не могут заразить человека. В этом выражается явление узкой специфичности этих видов паразитов.
Паразиты крови. Значительное число видов жгутиконосцев являются паразитами крови. Их среда обитания - жидкая часть (плазма) крови. Кровяные жгутиконосцы относятся к роду трипаносом (Trypanosoma). У человека паразитирует Trypanosoma gambiense, возбудитель тяжелого заболевания -сонной болезни. Это заболевание широко распространено в Экваториальной Африке. На ранних стадиях заболевания трипаносома живет в плазме крови, на поздних стадиях она проникает в спинномозговую жидкость.
Строение трипаносом чрезвычайно своеобразно и характерно (рис, 58). Тело их продолговатое, суженное на обоих концах и сплющенное в одной плоскости. Длина равняется 15-30 мк. От переднего конца вперед выдается короткий жгутик. Он продолжается вдоль почти всего тела и соединяется с его краем тонкой протоплазматической перепонкой, образуя, как и у трихомонад,ундулирующую мембрану. Несколько не доходя до заднего конца тела, базальная часть жгутика входит внутрь цитоплазмы. У основания его расположено базальное зерно и особое, резко окрашивающееся различными красками довольно крупное тельце, которое носит названиекинетопласта. Наблюдения за живыми трипаносомами в крови показывают, что движение их осуществляется благодаря энергичным волнообразным движениям ундулирующей мембраны. Ядро одно, несколько сдвинуто с середины тела кзади. В цитоплазме содержатся резервные питательные вещества в виде разной величины гранул. Питание чистоосмотическое (сапрофитное). Размножение обычно осуществляется путем продольного деления.
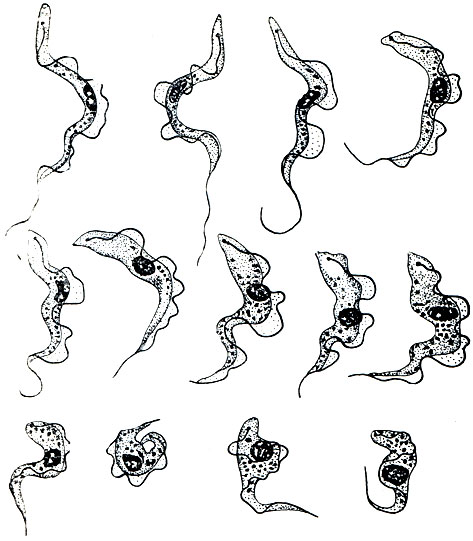
Рис. 58. Различные формы жгутиконосца Trypanosoma gambiense - возбудителя сонной болезни человека
Заболевание человека сонной болезнью при отсутствии лечения протекает очень тяжело. Первый период заболевания характеризуется явлениями лихорадки. Во втором периоде (когда трипаносомы проникают в спинномозговую жидкость) наблюдаются нервно-психические явления, сонливость, резкое истощение, которые приводят к смерти. В настоящее время существуют специфические лекарственные вещества (например, Германии), которые излечивают это тяжелое заболевание. В XIX в., когда терапия сонной болезни еще не была разработана и неизвестны были специфические лекарственные вещества, сонная болезнь представляла страшный бич Африки, от нее погибало огромное количество людей.
Какими путями происходит заражение человека трипаносомами, как проникают они в кровяное русло? Этому вопросу были посвящены многочисленные исследования, в результате которых удалось выяснить весь жизненный цикл возбудителя сонной болезни. Оказалось, что переносчиком трипаносомы является кровососущая муха цеце (Glossina palpalis), широко распространенная в Африке (рис. 59).

Рис. 59. Переносчик сонной болезни кровососущая муха Glossina palpalis (цеце): 1 - со спинной стороны; 2 - сбоку, насосавшаяся крови
Когда муха насосется крови от больного сонной болезнью, то в кишечнике ее трипаносомы не погибают. Напротив, они энергично размножаются, несколько меняя при этом свою форму. Образуется так называемаякритидиальная стадия, которая отличается от трипаносом крови тем, что кинетопласт у нее сдвигается несколько вперед и оказывается расположенным на уровне ядра. После периода размножения трипаносомы вновь приобретают типичную форму и активно проникают в слюнные железы и хоботок мухи. Эти "метациклические формы" являются инвазионными, т. е. способными заражать. Если цеце с метациклическими трипаносомами укусит человека, то трипаносомы проникают в кровяное русло, где и размножаются, вызывая заболевание сонной болезнью.
Trypanosoma gambiense паразитирует, кроме человека, в крови антилоп. Однако, в отличие от человека, антилопы не страдают от трипаносом, не обнаруживают никаких симптомов заболевания. В природных условиях антилопы являются постоянным источником заражения человека, так как мухи цеце в одинаковой степени нападают на человека и на антилоп"Сонная болезнь - это типичный пример трансмиссивного (передающегося через переносчика) заболевания с природной очаговостью (по терминологии акад. Е. Н. Павловского). Борьба с такими заболеваниями особенно затруднительна, так как не только больной человек является источником распространения заболевания, но, кроме того, существует природный очаг возбудителя, представляющий постоянную угрозу как источник инфекции.
Трипаносомы паразитируют, кроме человека, в самых разнообразных позвоночных животных (в рыбах, амфибиях, пресмыкающихся, птицах и млекопитающих). Ряд видов трипаносом паразитирует и в домашних животных, вызывая подчас очень тяжелые заболевания.
Для крупного рогатого скота в Африке одним из наиболее опасных и широко распространенных заболеваний является болезнь, называемаянагана. Она вызывается одним из видов трипаносом (Trypanosoma brucei), которая также передается кровососущими мухами из рода Glossina.
В Южной Азии не менее распространена другая болезнь -сурра, вызываемая трипаносомой Trypanosoma brucei. Переносчики ее - насекомые из отряда двукрылых - слепни. Однако в слепнях, в отличие от Glossina, трипаносомы не размножаются и никаких морфологических изменений не претерпевают. Слепни являются чисто механическими передатчиками инвазионного начала, так как трипаносомы, находясь у них в хоботке, сохраняют жизнеспособность несколько часов.
В Советском Союзе трипаносомные заболевания рогатого скота не играют сколько-нибудь заметной роли. Гораздо более распространены они в южных районах СССР у верблюдов и лошадей. Возбудитель трипаносомозов верблюдов, по-видимому, тождествен Trypanosoma evansi и передается слепнями. У лошадей трипаносомы вызываютслучную болезнь (возбудитель - Trypanosoma equiperdum). Возбудитель этого заболевания не передается переносчиками, а переходит от одного животного к другому через слизистые половых путей в момент случки.
К семейству трипаносомид относятся также представители рода леашманий (Leishmania), два вида которых являются возбудителями тяжелых заболеваний человека. Leishmania tropica вызывает кожное заболевание, известное под именемлейшманиоза илипендинской язвы. Эта болезнь широко распространена в Южной Азии, Северной Африке, в некоторых районах Южной Европы (Италия). В дореволюционное время пендинская язва часто наблюдалась в России в Средней Азии (Туркестан, Узбекистан). В настоящее время благодаря большой лечебной и профилактической работе, проведенной советскими органами здравоохранения, заболевание это сведено до минимума и встречается в виде редких единичных случаев.
Заболевание начинается с того, что на коже (чаще всего на открытых частях тела - на руках, лице) появляется узелок, увеличивающийся до размеров ореха, а затем на этом месте возникает открытая язва. Она очень долго не заживает (1 - 2 года), затем зарубцовывается. Возбудитель заболевания - Leishmania tropica - локализуется в подкожной клетчатке, преимущественно в соединительнотканых клетках (рис. 60), частью - в межклеточных промежутках. Возбудитель представляет собой мелкие овальные тельца длиной 4-7 мк, шириной 3-4 мк. Они в большом количестве заполняют клетки хозяина. Каждая клетка паразита имеет одно ядро и окрашивающееся тельце, которое является не чем иным, как кинетонуклеусом, столь характерным для трипаносом. От кинетопласта отходит базальная часть жгута. Однако ундулирующая мембрана и свободная часть жгутика не выражены. Это вполне понятно, так как при внутриклеточном паразитизме лейшмании, в отличие от трипаносом, неподвижны. Однако если их выделить в культуру (они легко культивируются на питательных средах), то у лейшмании развиваются жгутики и они активно плавают (рис. 61).

Рис. 60. Возбудитель кожного лейшманиоза (пендинской язвы) Leishmania tropica: 1 - группа паразитов, выпавшая из клеток хозяина; 2 - клетка с многочисленными паразитами
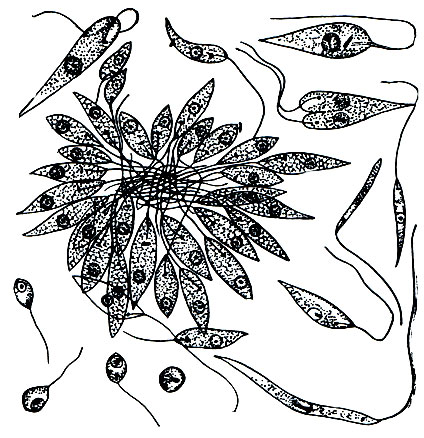
Рис. 61. Возбудитель висцерального лейшманиоза (кала-азара) Leishmania donovani в культуре. У жгутиконосца развиты жгутики, отсутствующие при паразитировании его в тканях хозяина
Каким же путем происходит заражение лейшманиями? В настоящее время можно считать доказанным, что переносчиками лейшмании являются москиты - очень мелкие кровососущие насекомые из отряда двукрылых (рис. 62), нападающие на человека преимущественно в ночное время. Москиты широко распространены в теплых странах и многочисленны у нас в Крыму, на Кавказе, в Средней Азии. Личинки москитов развиваются во влажной почве. Экспериментально было выяснено, что москиты заражаются лейшманиями, которые в кишечнике у них переходят в жгутиковую стадию (так называемая "лептомонадная форма" - название, обязанное сходству со жгутиконосцами рода Leptomonas). Здесь же происходит размножение лейшманий. Затем часть их перемещается в самый передний отдел кишечника и при укусе вводится в кожу жертвы.

Рис. 62. Переносчик лейшманиозов москит Phlebotomus pappatasii. Самка, увеличенная в 13 раз
Многолетние исследования советских ученых (главным образом проф. Н. Латышева и его сотрудников) показали, что пендинская язва также является трансмиссивным заболеванием с природной очаговостью. Лейшманиями могут заражаться грызуны, а также собаки. Естественный цикл паразита обычно завершается грызунами и обитающими в их же норах москитами, минуя человека. Достаточно человеку попасть в такой естественный очаг лейшманиоза, чтобы он заразился пендинской язвой вне зависимости от наличия таковой у окружающих его людей.
Вторая болезнь человека, вызываемая лейшманиями, -висцеральный (внутренностный)лейшманиоз, иликала-азар. Возбудитель его Leishmania donovani морфологически неотличим от описанной выше Leishmania tropica. Переносчиками и здесь являются москиты. Кала-азар также распространен в Южной Азии, Северной Африке, Малой Азии. В Советском Союзе известен в Средней Азии (Туркменской ССР). В Европе встречается в Южной Италии, Греции. Это очень тяжелое заболевание, проявляющееся в лихорадке, малокровии, общем истощении и гипертрофии печени и селезенки, при отсутствии лечения обычно приводящее к смертельному исходу. Возбудитель поражает различные внутренние органы, особенно часто встречается в эндотелии кровеносных сосудов, в селезенке, печени, костном мозгу, лимфатических железах. Это заболевание чаще поражает детей, чем взрослых.
Кроме человека, носителями лейшманий, вызывающих кала-азар, могут быть собаки, которые в распространении этого заболевания играют, по-видимому, немаловажную роль.
Паразитические жгутиконосцы земноводных. В заднем отделе кишечника земноводных обитают многочисленные виды паразитических простейших - жгутиконосцев и инфузорий, легко доступных для наблюдения. Из жгутиконосцев особенно богато представлены разные виды семейства опалинид (Opalinidae). В заднем отделе кишечника лягушек в огромных количествах встречаются разные виды рода опалин (Opalina). Это очень крупные простейшие, достигающие 1 мм. У наиболее обычной в наших травяных лягушках О. ranarum, как и у других опалин, тело плоское, листовидное. Оно не вполне симметрично: косо срезано спереди и закруглено сзади (рис. 63). Поверхность тела покрыта многочисленными жгутиками, количество которых измеряется многими тысячами. Они расположены продольными и несколько S-образно изгибающимися рядами. Хорошо разграничены эктоплазма и эндоплазма. В эндоплазме разбросано несколько десятков небольших ядер. Здесь мы встречаемся с примером многоядерного простейшего. Сократительных вакуолей у опалин нет.
Среди опалин нередко встречаются делящиеся особи. Борозда деления при этом проходит косо (рис. 63), что не позволяет это деление рассматривать ни как продольное, ни как поперечное.
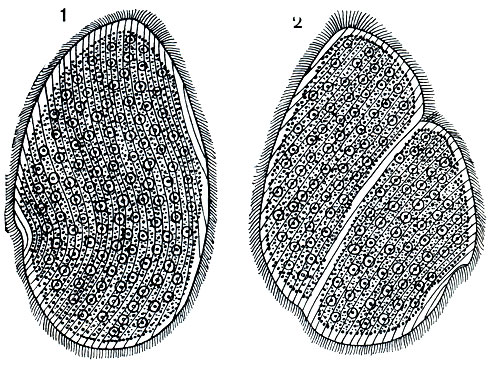
Рис. 63. Паразитический жгутиконосец из задней кишки лягушки Opalina ranarum. Видно большое количество жгутиков и ядер. 1 - неделящаяся особь; 2 - деление, борозда проходит косо по отношению к длинной оси простейшего
До недавнего времени опалин причисляли к инфузориям в качестве самостоятельного подкласса. Основанием для этого служило большое количество ресничек (жгутиков), которые целиком покрывают все тело сплошными рядами (см. ниже об инфузория,). Однако у опалин отсутствуют основные черты, характерные для инфузорий: ядерный дуализм и половой процесс типа конъюгации. Это заставило выделить их из инфузорий и рассматривать как своеобразное многожгутиковое простейшее из класса жгутиконосцев.
У опалин нет ротового отверстия и питание совершается осмотическим путем.
Как осуществляется заражение новых особей хозяина, как заражаются ими головастики? Основные этапы жизненного цикла хозяина (лягушки) и паразита (опалин) очень точно "синхронизированы", что обеспечивает заражение.
В лягушках, пойманных ранней весной в момент икрометания, встречаются очень мелкие особи опалин, образующиеся в результате многократных делений. Это предцистные формы. Они инцистируются и выходят из лягушки, падая на дно водоема. Там они лежат до тех пор, пока не будут проглочены головастиками. В их кишечнике из цист вылупляются мелкие опалины, которые путем деления дают начало еще более мелким одноядерным клеткам, являющимся половыми (гаметами). Эти клетки попарно сливаются (половой процесс -копуляция), причем происходит слияние не только цитоплазмы, но и ядер. Из образовавшихся зигот в дальнейшем развиваются многоядерные опалины. Интересно отметить, что половой процесс опалин совершается только на одном этапе цикла - в головастиках. Полового процесса у опалин в теле лягушек никогда не происходит, и размножаются они бесполым путем.
Жгутиконосцы, обитающие в кишечнике термитов и тараканов. Термиты - это большая группа (отряд) преимущественно тропических насекомых, характерных своим "общественным" образом жизни. На территории Советского Союза термиты встречаются в Средней Азии и в окрестностях Одессы. Эти насекомые, образующие большие колонии и строящие сложные гнезда - термитники, питаются преимущественно растительной пищей, в том числе древесиной, поглощая ее в больших количествах. В кишечнике термитов, а именно в задней кишке, образующей большое расширение, живет огромное количество жгутиконосцев. Видовой состав их исключительно разнообразен. В настоящее время описано свыше 200 видов жгутиконосцев, обитающих в кишечнике термитов. При этом нужно иметь в виду, что в паразитологическом отношении изучено не более1/3 общего числа видов термитов. Учитывая, что некоторые паразитические простейшие обладают довольно узкой специфичностью, можно думать, что число видов их значительно больше известного в настоящее время.
Жгутиконосцы из кишечника термитов относятся к двум отрядам: Polymastigina и Hypermastigina. Это наиболее сложно устроенные среди жгутиконосцев формы. Их паразитический образ жизни не только не привел к упрощению организации, но, напротив, связан с ее усложнением. В этом сходство их с инфузориями, населяющими кишечный тракт копытных (рис. 64). Количество простейших в кишечнике термитов бывает настолько велико, что оно составляет иногда до1/3 веса тела насекомого!
На рисунке 64 изображены некоторые представители жгутиконосцев, обитающих в кишечнике термитов. Для многих из них характерно большое количество жгутиков, которые или сосредоточены на переднем конце тела, или - в других случаях - занимают значительную часть его поверхности. Жгутики могут быть расположены продольными рядами, по спирали (относительно продольной оси тела) или поперечными рядами. Иногда жгутики собраны пучками на переднем конце (рис. 64). Размеры жгутиконосцев различны, они варьируют от 30-40 мк до 0,5 мм и более.

Рис. 64. Различные жгутиконосцы из кишечника термитов. А - Teratonympha mirabilis; Б - Spirotrichonympha flagellata; В - Coronympha octonaria; Г - Calonympha grassi; Д - Trichonympha turkestanica; E - Rhynchonympha tarda, 1 - ядра; 2 - аксостили
Ядерный аппарат у разных представителей устроен неодинаково. Некоторые виды (рис. 64,Д) имеют одно крупное ядро. Другие являются многоядерными. Иногда число ядер невелико (рис. 64,Б), и они располагаются венчиком на переднем конце тела. Иногда же число ядер достигает нескольких десятков (рис. 64,Г), и они сосредоточены в передней трети тела.
Характерной особенностью строения некоторых жгутиконосцев, обитающих в кишечнике термитов, является наличие у них опорных образований -аксостилей. Пример таких эластических скелетных структур мы уже видели у трихомонад и лямблий. Часто имеется не один, а целый пучок аксостилей (рис. 64,Г).
В цитоплазме жгутиконосцев, обитающих в кишечнике термитов, обычно имеется большое количество пищевых включений. Это кусочки растительных тканей разных размеров. Заглатывание пищи происходит различными способами. Ротовое отверстие у большинства видов отсутствует. Часть их цитоплазмы не имеет резко очерченной пелликулы, а сохраняет способность образовывать выросты, напоминающие псевдоподии. При помощи таких выростов жгутиконосец заглатывает частицы растительных тканей, присутствующие всегда в большом количестве в кишечнике термита. На рисунке 65 изображены последовательные стадии заглатывания жгутиконосцем из рода трихонимфа (Trichonympha) довольно крупного кусочка древесины. Захват пищи осуществляется и другим путем. Для заглатывания мелких частиц на заднем конце жгутиконосца образуется временное впячивание, в которое втягиваются кусочки пищи. Впячивание замыкается сзади, и пищевые частицы оказываются лежащими в цитоплазме простейшего.

Рис. 65. Жгутиконосец из кишечника термитов Trichonympha sp., заглатывающий кусочек древесины
Разные стадии жизненного цикла возбудителей человеческой малярии в эритроцитах крови человека и в слюнных железах комара-переносчика Anopheles с препаратов, окрашенных по Гимза-Романовскому
Установлено, что взаимоотношения между термитами и населяющими их кишечник жгутиконосцами - это явление тесногосимбиоза: жгутиконосцы могут существовать только в кишечнике термитов, содержимое которого является для них средой обитания и источником пищи. Но и термиты не могут существовать без жгутиконосцев. Пищей для термитов служит почти исключительно клетчатка (древесина). Переваривание заглоченной пищи происходит в основном в заднем отделе кишечника, заселенном жгутиконосцами. В кишечнике самих термитов не выделяется ферментов, расщепляющих клетчатку. Напротив, в теле симбиотических жгутиконосцев исследованиями Хангейта обнаружен фермент целлюлаза, расщепляющий клетчатку. Жгутиконосцы переводят клетчатку в растворимое состояние, используя при этом лишь часть переработанного вещества. Таким образом, питание термитов в значительной мере осуществляется при участии симбиотических простейших. Это доказывается прямыми опытами, проведеннымиКливлендом и подтвержденными многими исследователями. Оказывается, довольно простым методом можно освободить термитов от жгутиконосцев, не повреждая самих насекомых. Если их поместить в чистый кислород при давлении в 3 атмосферы, все простейшие вскоре погибают, тогда как термиты остаются живыми. После освобождения насекомых от простейших Кливленд помещал их в нормальные условия с обильным количеством древесной пищи. В таких же условиях жили и нормальные термиты, т. е. содержащие простейших в кишечнике. Термиты со жгутиконосцами жили в условиях опыта в стеклянных сосудах долгое время, до 1 -11/2 лет, пока не наступала естественная смерть их от старости. Напротив, насекомые, лишенные жгутиконосцев, выживали не более 10-14 дней и потом неизбежно погибали. При этом они энергично поглощали пищу, но она не могла у них перевариваться. Стоило искусственно заразить таких "стерильных" термитов жгутиконосцами, как они приобретали способность переваривать пищу и жить продолжительное время.
Мы уже говорили, что каждая особь термита имеет в кишечнике жгутиконосцев. Возникает вопрос: какими путями происходит проникновение в кишечник молодых личинок их симбионтов - жгутиконосцев? У термитов существует особый инстинкт слизывания друг у друга жидких экскрементов, выступающих в форме капельки из анального отверстия. Термит, проглатывающий содержимое задней кишки особи того же вида, получает некоторое количество усвояемой пищи, переведенной в растворенное состояние благодаря деятельности жгутиконосцев. С другой стороны, проглатывая жгутиконосцев, он вводит их в свой кишечник, где они продолжают жить и размножаться. Так же заражаются и молодые личинки.
В колонии термитов лишены жгутиконосцев лишь половозрелые взрослые особи. В термитнике обычно имеется одна самка ("царица"), непрерывно откладывающая яйца. Эта самка не питается древесиной. Ее кормят "рабочие", которые представляют собой недоразвитых в половом отношении особей. Пищей служит отрыжка из переднего отдела кишечника " содержащая легкоусвояемые вещества.
Фауна жгутиконосцев развивается, кроме термитов, в кишечнике растительноядных тараканов рода Cryptocercus.
Рост и развитие термитов, как и других насекомых, сопровождается периодической линькой, в процессе которой сбрасывается тонкая хитиновая выстилка заднего отдела кишечника. При этом у большинства видов термитов выходят наружу и жгутиконосцы, и насекомое после линьки оказывается лишенным простейших. Очень скоро они вновь появляются, так как слинявшее насекомое начинает энергично слизывать жидкие экскременты окружающих особей. У тараканов Cryptocercus во время линьки жгутиконосцы заползают в просвет между отделяемой внутренней выстилкой и эпителиальной стенкой кишки, откуда по окончании линьки вновь выходят и заполняют просвет кишечника.
У жгутиконосцев, обитающих в кишечнике термитов, известно толькобесполое размножение путем деления. Половой процесс у них отсутствует. Иное делю жгутиконосцы из кишечника таракана Cryptocercus. Работами Кливленда установлено наличие у них полового процесса по типу копуляции, который протекает строго периодически. Оказалось, что половые процессы жгутиконосцев находятся в строгом соответствии с линькой личинок. Как известно, рост насекомых на личиночных стадиях сопровождается периодическим сбрасыванием хитинового покрова, так как последний может растягиваться лишь до определенного предела, а затем, после линьки, заменяется новым хитиновым покровом. Процесслиньки - очень сложное явление. Наступление его связано с действием гормонов, которые выделяются некоторыми эндокринными железами насекомого и обусловливают отслаивание старой хитиновой кутикулы. У разных видов жгутиконосцев половой процесс происходит строго закономерно в разные сроки подготовительного к линьке периода или же вскоре (через 1-2 дня) после завершения линьки. Для каждого вида жгутиконосцев соотношение срока наступления полового процесса и линьки является строго определенным. Очевидно, выделяемые хозяином гормоны линьки воздействуют не только на насекомое, но и на обитателей его кишечника, стимулируя половое размножение. У взрослых тараканов, которые не растут и не линяют, половых процессов у населяющих кишечник жгутиконосцев не происходит.
Жгутиконосцы - паразиты растений. Некоторые жгутиконосцы приспособились к паразитированию в растительных тканях. Хозяевами их являются растения с млечным соком. Это виды семейств молочайных, сложноцветных, маковых, ластовниковых и др. Жгутиконосцы-паразиты растений относятся к роду Leptomonas, принадлежащему к семейству трипаносомид, с некоторыми представителями которых мы познакомились выше. Виды этого рода имеют одно ядро, один жгутик и кинетопласт, расположенный впереди ядра (рис. 66).
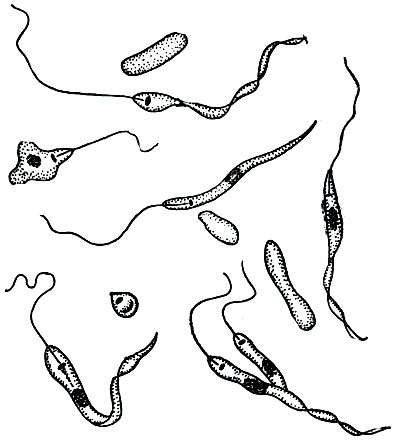
Рис. 66. Жгутиконосец Leptomonas davidi из млечного сока молочая
Живут Leptomonas в сосудах, наполненных млечным соком, в межклетниках, в вакуолях отдельных клеток. Нередко эти простейшие развиваются в растительных тканях в огромных количествах.
Массовое развитие жгутиконосцев наносит растениям большой вред, вызывает разрушение хлорофилла, опадение листьев и в конечном счете гибель растения. Жгутиконосцы-паразиты растений распространены преимущественно в тропических и субтропических странах, в умеренной зоне встречаются реже. Очень большой вред паразитические жгутиконосцы наносят в некоторых странах плантациям кофе. Особенно широко это заболевание кофе распространено в Гвиане (Южная Америка). Оно носит название некроз флоэмы, сопровождается пожелтением и опадением листвы. При сильном заражении кофейные деревца погибают в течение 3-12 месяцев.
Перенос возбудителя и заражение растений осуществляется насекомыми. Обычно переносчиками являются растительноядные клопы, сосущие СОКИ растений.
Обзор важнейших представителей класса жгутиконосцев показал, насколько разнообразны эти организмы. Этот класс стоит как бы на границе растительного и животного мира, и многие представители его совмещают столь различные типы обмена веществ, как аутотрофный и гетеротрофный.
Эволюция жгутиконосцев шла в различных направлениях. Многие ветви их развития привели к образованию планктонных форм, примером которых являются панцирные жгутиконосцы. Особенно широкое развитие получил у жгутиконосцев паразитизм. Многократный переход к паразитизму осуществлялся в разных линиях их филогенетического развития. В ряде случаев на основе паразитизма возникли очень тесные симбиотические взаимоотношения, примером которых могут служить жгутиконосцы кишечника термитов и некоторых тараканов.
Интересным путем эволюции жгутиконосцев является переход к колониальным формам организации. Колонии их очень разнообразны и достигают у высших представителей (например, Volvox) очень сложной дифференцировки, выходящей уже за пределы одноклеточности. Колонию вольвокса, может быть, правильнее рассматривать как многоклеточный организм, поскольку, как мы видели, не все клетки колонии равноценны. Возможно, что колониальность имела большое значение в эволюции органического мира и явилась переходным этапом к многоклеточным животным.