

Некоторые этапы эволюции животных
Эволюционное развитие живых существ следует рассматривать как целостный процесс развития живого населения биосферы от начальных, примитивных форм до современных, наиболее совершенных. Это относится в равной мере к морфологическим и биохимическим структурам и к физиологическим процессам.
Один из интереснейших вопросов в эволюции организмов - происхождение многоклеточности. Никто не подвергает сомнению, что многоклеточные организмы произошли от одноклеточных, и большинство ученых сходится на том, что родоначальниками многоклеточных были колониальные простейшие.
Биологический смысл возникающейколониальности* заключается в ее защитной роли от врагов и от факторов абиотической среды.
* (Термин колониальность в зоологии употребляется в двух смыслах: в данном случае имеется в виду морфологическое объединение нескольких особей общими образованиями, а иногда путем срастания в индивидуум высшего порядка. Однако колониальностью именуют и сосуществование многих особей скоплениями или колониями. В этом смысле говорят о колониях птиц, о гнездах общественных насекомых и т. п.)
Среди высших многоклеточных колониальность очень ярко выражена у мшанок и оболочниковых. Впрочем, следует отметить, что колониальность отсутствует у многих больших групп животных, например у моллюсков, членистоногих, аннелид, иглокожих, позвоночных и некоторых других, но очень сильно выражена у простейших и кишечнополостных, т. е. в тех группах, которые стоят у истоков многоклеточности. Особенно интересны, конечно, те колониальные организмы, у которых наблюдается морфологическое и функциональное разделение между особями колонии по основным жизненным функциям, как движение, питание, размножение, защита. На низшей ступени развития колониальности находятся такие формы, как Pandorina или Eudorina среди жгутиковых, между сочленами колонии которых не наблюдается различий. В данном случае колониальный организм - индивидуум второго порядка - еще не подчиняет себе отдельные организмы, входящие в состав колонии. С усложнением колониальности происходит все большее подчинение отдельной особи целому - индивидууму второго порядка, а "разделение труда" между особями идет по линии функций движения и размножения, а в дальнейшем и питания. Развитию колонпальности сопутствуют явления полиморфизма, которые наблюдаются также и при формах колониального существования насекомых. Совместное существование обычно ведет к морфологическому и функциональному разделению между особями.
Явлениеполиморфизма (многоформия), так же как и колониальность, возникает независимо в разных группах животных. Даже в пределах одного класса (например, термиты и муравьи среди насекомых) явления полиморфизма могут возникать совершенно независимо и давать при этом поразительное конвергентное сходство. Полиморфизм наиболее ярко в животном царстве выражен у кишечнополостных, хотя природа их совсем другая, чем у насекомых. Эта широко развитая у простейших и кишечнополостных способность к образованию колоний дает основание думать, что именно таким образом сформировалась многоклеточность, а многоядерность простейших следует рассматривать как усложнение строения ядра, не ведущее к образованию многоклеточности. При этом большинство многоядерных простейших характерно для паразитических форм (Sporozoa), организация которых не может быть положена в основу филогенетических построений, а должна рассматриваться как результат крайне специфических условий существования (см. раздел "Простейшие").
С формированием многоклеточности получил развитие эмбриональный процесс - процесс формирования из оплодотворенного яйца взрослой формы.
Прежде чем перейти к некоторым моментам, связанным с эмбриональным процессом, следует затронуть одно из наиболее замечательных обобщений, связанных с индивидуальным развитием животных, так называемыйбиогенетический закон, или, как его сейчас обычно именуют,правило рекапитуляции. Это правило глубоко проникло в современные эмбриологию и сравнительную анатомию, так же как принципы монофилии и дивергентной эволюции. Биогенетический закон связывается с именами двух крупнейших последователей Ч. Дарвина - Э. Геккеля и Ф. Мюллера, хотя еще и до них некоторые зоологи близко подходили к этому обобщению (Меккель, Бэр), в том числе и Дарвин. В основном это обобщение заключается в том, что в онтогенетическом развитии организмов мы обнаруживаем в некоторой последовательности отражение этапов, пройденных в прошлом, или, как иногда говорили в краткой форме, "онтогенез повторяет филогенез".
Конечно, в такой категорической форме сейчас биогенетический закон уже не принимают. В развитии организма часто смещается последовательность появления признаков эволюционного прошлого.
Наиболее характерная особенность процесса совместного овладения и другими живыми существами жизненным пространством - это переход от менее совершенных форм к более совершенным, от менее сложных к более сложным, от менее дифференцированных к более дифференцированным, от менее специализированных к более специализированным, при этом от более разнообразных к менее разнообразным, но более усложненным и специализированным в соответствии с дарвиновским принципом дивергентной эволюции. В отдельных случаях, однако, при изменении биологии организма и его организации, при смене среды обитания можно наблюдать на отдельных системах органов регрессивные изменения, приводящие к значительной редукции, а иногда и к полному исчезновению тех или иных особенностей организации. Параллельно с такими процессами наблюдается обычно сильное развитие или появление других систем органов, которые и сопутствуют указанной редукции. Прежде всего эти процессы сопутствовали выходу из водной среды в воздушную, переходу к существованию в глубинах океана, в пещерах, переходу к паразитическому существованию, изменениям в характере питания и т. д.
В эволюции животных изменения сходного типа имеют место при увеличении размеров тела, при повышении уровня организации в эволюционном процессе. Так, нематоды и членистоногие утеряли ресничный эпителий в связи с кутикуляризацией покровов, а выход многих животных в воздушную среду привел к редукции дыхательного аппарата, свойственного водным организмам (жабры).
При переходе животных к малоподвижному или неподвижному образу жизни и к паразитизму происходят значительные изменения в отдельных органах и целых системах, причем эти изменения могут идти в диаметрально противоположных направлениях: у свободноживущих организмов переход к неподвижному существованию сопровождается сильным развитием механических или химических защитных образований (скелеты губок, кишечнополостных и мшанок, раковины моллюсков и плеченогих, домики усоно-гих ракообразных, стрекательный аппарат кишечнополостных, ядовитые или отталкивающие выделения кожных желез, слизь и т. д.). Но наряду с этим редукции подвергаются мускулатура, нервная система и органы чувств, изменяются способы питания и захвата добычи. У паразитов скелетные образования отсутствуют, но развиваются различные органы прикрепления к органам хозяина (присоски, крючки, хоботки). В обоих случаях имеет место сильнейшее увеличение плодовитости, ослабление роли особей мужского пола и часто развитие гермафродитизма (усоногие ракообразные). При паразитизме часто наблюдается полная редукция кишечника (цестоды, колючеголовые), что до недавнего времени не было известно у свободноживущих животных. Совершенно парадоксальным случаем оказалось установленное А. В. Ивановым полное исчезновение кишечника у глубоководной группы погонофор, что пока не находит себе объяснения.
Весьма интересно, что часто изменения организации, сопутствующие смене среды обитания, даже у родственных форм происходят не однообразно, а иногда идут диаметрально противоположными путями. Например, у одних глубоководных рыб наблюдается темная, почти черная окраска, а другие становятся бесцветными и иногда прозрачными. У одних глубоководных рыб наблюдаются гипертрофированные, иногда стебельчатые глаза, а у других - полная редукция органов зрения. При выходе из водной среды в воздушную у большинства животных выработались рычажные органы движения - ноги (членистоногие, позвоночные), однако многоножки и змеи имеют длинное тело и передвигаются изгибанием тела, что чаще свойственно водным организмам.
Крайнюю форму частичной или полной редукции многих органов можно наблюдать у паразитических усоногих ракообразных (Sacculina, Dendrogaster и др.) и брюхоногих моллюсков (Enteroxenos и Parenteroxenos).
Все эти бесчисленные случаи редукции органов нельзя рассматривать как процессы регресса вообще. Вернее считать их формами выработки узкоспециализированных черт организации как приспособления к аномальным условиям существования и видеть в них яркое выражение огромной способности живых существ приспосабливаться к самым различным условиям существования и к расширению своего жизненного ареала. Таким образом, все это следует рассматривать как формы приспособления, а не явления общего регресса.
Возвращаясь к общей тенденции эволюционного процесса - переходу от менее сложных, но более разнообразных структур к немногим (иногда всего лишь одной), но высоко и многообразно специализированным, остановимся на ряде примеров. Хорошим примером может быть эволюция форм движения. Простейшие обладают весьма разнообразными формами движения. Саркодовые (особенно корненожки) имеют псевдоподиальное движение, которое осуществляется по гидравлическому принципу - эндоплазма устремляется в тот или иной участок периферии и "натягивает" в более плотной и упругой эктоплазме вырост -псевдоподию.
Движение жгутиконосцев обеспечивается винтообразным биением жгутиков, а инфузорий - многочисленными ресничками. В противоположность жгутикам реснички бьют в одной плоскости, но сами они служат, помимо плавания, разным целям: окружающие ротовое отверстие реснички создают сложный ток воды, загоняющий пищевые частицы в глотку. Реснички могут слипаться в "кисточки" -цирры и на брюшной стороне брюхоресничных инфузорий имитируют конечности. На этих циррах инфузории могут быстро "бегать" по субстрату. Соединяясь в продольные ряды, реснички превращаются в мембраны, способные к волнообразным колебаниям. У трипанозом вдоль всего тела идут также мембраны, по краю их проходит основание жгута и мембрана все время находится в состоянии волновых колебаний. Грегарины движутся реактивным способом, отталкиваясь от стекающей с заднего конца тела слизи. Простейшим свойственны также многообразные формы парения в воде при помощи радиально расходящихся скелетных игл или псевдоподий.
Ресничное движение свойственно также бесчисленным личинкам донных животных (черви, моллюски, иглокожие и др.) и самым мелким многоклеточным, сходным по размерам с простейшими (коловратки и некоторые турбеллярии).
Ресничное и жгутиковое движения имеют еще одну особенность - двигающиеся таким образом организмы при плавании совершают вращательное движение вокруг собственной оси (кроме турбеллярий) и, кроме того, двигаются не прямолинейно, а по винтовой линии. Таким способом осуществляется у этих микроскопических существ, имеющих удельный вес, близкий к воде, двойное двигательное вбуравливание в водную среду. Этот способ движения в воде весьма эффективен. Относительная скорость инфузорий (отношение скорости проходимого в секунду пути к длине тела), перенесенная на размеры тела человека, составила бы скорость спринтера.
Таким образом, у простейших мы наблюдаем всевозможные способы движения, кроме полета. У кишечнополостных способностью к свободному движению обладают только медузы, сифонофоры и ктенофоры. Ктенофоры движутся при помощи ресничного эпителия, но движение их крайне медленное. Ресничное движение эффективно только при малых величинах тела, измеряемых долями миллиметра. С увеличением размеров тела ресничное и жгутиковое движения становятся неэффективными, так как объем тела возрастает гораздо быстрее (в кубе), чем его поверхность (к квадрате). В дальнейшем у червей сформировалась новая форма движения - изгибанием тела, а в связи с этим у них развилась мощная двигательная мускулатура. У кишечнополостных мускулатура была совершенно недостаточной для выработки новой формы движения. 95% кишечнополостных ведут неподвижное существование, защищаясь от врагов мощным скелетом и стрекательным аппаратом. Еще более ярко это сказалось на губках, совсем лишенных мускулатуры.
Движение медуз и сифонофор совершается реактивным способом - колокол медузы сжимается, выталкивает из-под себя воду, и медуза получает реактивный двигательный толчок верхней стороной колокола вперед. Для такого движения достаточно небольшой двигательной мускулатуры, которая составляет едва 1 - 2% от объема тела. Только у червей начинает накапливаться двигательная мускулатура в виде кожно-мускульного мешка. У немертин и высших червей она достигает наибольшего развития.
У кишечнополостных между эктодермой и энтодермой уже создается мышечный слой (первоначально в очень примитивной форме) в виде слоев сократительных отростков эпителиально-мышечных клеток, образующих два взаимно перпендикулярных волокна - клетки эктодермы образуют систему продольных по оси тела волокон, а клетки энтодермы - кольцевых. Эта система волокон как бы имитирует кожно-мускульный мешок червей, но не осуществляет изгибательных движений, а осуществляет только сократительные - по основной оси тела. Кожно-мускульный мешок червей состоит из двух основных слоев мышц - кольцевого и продольного. Есть еще и другие мышцы: тело червей как бы набито мышцами, количество их достигает у некоторых червей 60-70% общего объема тела ( немертины, пиявки). Огромное количество низших и высших червей передвигается при помощи изгибаний тела. Таким образом, внутри группы кишечнополостных вместе с выработкой двустороннесимметричного плана строения сформировались мускульные формы животных, выработавшие изгибательный способ движения. Это были предки турбеллярий, использующие более мощную мускулатуру для изгибаний тела. Хотя изгибательное движение и доминирует у червей, но наряду с ним есть и некоторые другие формы движения.
Весьма сходные процессы, которые привели к тому же изгибательному движению, имели место также и в эволюции вторичноротых.
Масса низших хордовых и родственных им форм, ведших или малоподвижное, или неподвижное существование, дала начало рыбообразным с характерным для них изгибательным движением и мощной туловищной мускулатурой. В течение долгого периода эволюции позвоночных животных основной формой их передвижения было изгибательное.
В дальнейшем усложнение изгибательной формы движения наступило с выработкой посегментных складок -параподий, снабженных щетинками (хетами). Это был добавочный орган движения, если можно так сказать, чреватый последствиями, так как в дальнейшем из них сформировались конечности членистоногих. Полимерное строение кольчецов соответственно дало формы с большим числом ног ( ракообразные, паукообразные) .
С формированием рычажных конечностей, за немногими исключениями ( многоножки и змеи), вся двигательная функция перешла на ножные придатки. Путь формирования конечностей у позвоночных был несколько иным по сравнению с таковым у членистоногих. У членистоногих конечности сформировались уже в водной среде ( ракообразные), а у позвоночных- в процессе выхода в воздушную среду. Замечательно, что и у предков позвоночных плавники тоже сформировались из складок по бокам тела, но только в виде двухпоясов - плечевого и тазового - с двумя нарами конечностей. Столь же интересно, что первоначально плавники у предков членистоногих и позвоночных играли роль не основных органов движения, а только подсобных, но важно, что в обоих случаях рычажные конечности, при всей их чрезвычайной функциональной усложненности (полет),- единственный аппарат движения. Многообразие форм движения сменилось одной формой, однако в крайней степени усложненной и дифференцированной.
Подобный же эволюционный путь прошло большинство систем органов и функций. Остановимся еще только на одной, например на функции размножения. Трудно найти другую функцию у животных, которая давала бы столь же исключительное разнообразие. Прежде всего, конечно, следует указать на обычность у низших групп животных бесполого размножения, иногда полностью заменяющего половое, как, например, у многих отрядов жгутиковых и некоторых саркодовых. При этом формы бесполого размножения бесконечно разнообразны: от создания огромных колоний кишечнополостных или мшанок, при добавочных явлениях полиморфизма, до правильного чередованиягамогонии (полового) иагамогонии (бесполого) у фораминифер и споровиков.Бесполое размножение у простейших носит характер деления и почкования, простого и множественного, вплоть до сложных форм шизогонии. Сам половой процесс имеет двойную форму -копуляции и конъюгации, а кроме того,партеногенеза и оплодотворения. Столь же многообразны формы чередования полового и бесполого развития.
С повышением организации диапазон и разнообразие форм бесполого размножения уменьшились, среди первичноротых оно встречается еще у кольчатых червей, а из вторичноротых - только у оболочниковых. В конце концов, бесполое размножение уступает место половому.
Партеногенез, как исключение, встречается у всех групп членистоногих и позвоночных, и существуют виды насекомых и рептилий, у которых самцы совсем неизвестны и размножение совершается только партеногенетически.
У кишечнополостных впервые появляются нервная система и органы чувств (зрения и равновесия). Нервные клетки генетически связаны не только с эктодермой, как у всех прочих животных, кроме иглокожих, но и с энтодермой. Расположение частей тела и органов подчинено у кишечнополостных радиально-осевой симметрии.* Симметрией называется определенный геометрический порядок в расположении сходственных частей тела.
* (Симметрия-от греческих слов sym - вместе и metron - масса, в смысле часть тела.)
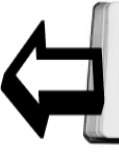
Элементами симметрии являются точка (центр), линия (ось) и плоскость. Прекрасный пример радиально-лучевой симметрии дают радиолярии (рис.3). Сходственные части тела расположены вокруг центра симметрии в радиальном направлении. Радиально-лучевая симметрия свойственна организмам, взвешенным в воде и имеющим со всех сторон одинаковую среду, в силу чего и реакция организма "одинакова во все стороны". Радиально-лучевая симметрия наилучшим образом соответствует биологии радиолярий. Радиально-лучевую симметрию мы находим также у колониальных фитомонадовых (вольвокс, эвдорина, пандорина и др.) и некоторых колоний многоклеточных, например у колониальной коловратки Conochilus.
Рис. 3. Различные типы симметрии у животных. А - радиально-лучевая симметрия; Б - радиально-осевая симметрия у кишечнополостных; В - двусторонняя симметрия
Однако радиально-лучевая симметрия некоторых простейших не является самой примитивной формой строения тела. В равной мере планктонное существование также нельзя считать самой примитивной биологической формой. Самые просто организованные формы саркодовых (Аmоеbina) имеют асимметричное строение, и, видимо, оно соответствует примитивным формам организации и поведения (псевдоподиальная форма движения и питания). Кроме того, можно думать, что все пелагические формы существования являются вторичными производными от придонных. Асимметричное строение свойственно и инфузориям, и жгутиковым. В частности, радиолярии обладают необыкновенным богатством планов симметрии своего скелета - радиально-осевой, как гомополярной, так и гетерополярной, двусторонней, двубоковой, с обычным отклонением всех этих типов симметрии в асимметрию. Следует отметить, что в подавляющем числе случаев при этом разные формы симметрии относятся только к скелету, что же касается протоплазмы, то она, как правило, имеет асимметричное расположение включений (ядро, пульсирующие и пищеварительные вакуоли и иные включения).
Кишечнополостным, как сидячим, так и пелагическим (медузы), свойственна радиально-осевая симметрия, при которой сходственные части расположены вокруг оси вращения, причем эта симметрия может быть самого различного порядка в зависимости от того, на какой угол следует повернуть тело животного, чтобы новое положение совпало с исходным. Таким образом, может получаться 4-, 6-, 8-лучевая симметрия и более, до симметрии порядка бесконечности. У радиолярий встречается радиально-осевая симметрия с одинаковыми полюсами, или, как говорят, гомополярная. У кишечнополостных - гетерополярная осевая симметрия: один полюс симметрии несет рот и щупальца (оральный), другой (аборальный) служит для прикрепления (стадия полипа), или у плавающих форм несет орган чувств (ктенофоры), или ничем не вооружен (медузы). У некоторых медуз на этой аборальной стороне образуется стебелек для прикрепления к подводным предметам (Lucernariida). Нарушение радиально-осевой симметрии возникает при уменьшении числа шупалец или изменении формы ротовой щели, пищевода и разветвлений пищеварительной системы. Количество щупалец может уменьшаться до одного (Моnоbrachium), и тогда их радиальное расположение сменяется двубоковым. Глотка может сплющиваться, и тогда тоже получается двубоковая симметрия, этому способствует и образование в глоткесифоноглифов (желобок вдоль глотки).
Наибольшее усложнение радиально-осевой симметрии наблюдается у ктенофор, где, помимо 8-лучевой симметрии, в расположении отдельных частей тела и органов наблюдается 4-лучевая и двубоковая симметрия. Это весьма существенный момент, так как большинство зоологов именно от ктенофорообразных предков выводит оба ствола высших животных, как первично-, так и вторичноротых.
Гетерополярная радиально-осевая симметрия вполне соответствует образу жизни кишечнополостных - неподвижному существованию в прикрепленном положении или медленному плаванию при помощи реактивного движения.
С другой стороны, от сложного типа радиально-осевой симметрии ктенофор можно перейти к двусторонней симметрии, или, как говорят, симметрии зеркального изображения, единственного плана симметрии трехслойных животных, симметрии быстрого движения, с выработкой переднего по движению конца тела, с центральным мозговым скоплением и основными органами чувств, спинной и брюшной, правой и левой сторонами тела. Однако живых или ископаемых свидетелей этого перехода мы не знаем. Здесь можно пользоваться только более или менее достоверными гипотезами.
В 1880 г. знаменитый эмбриолог А. Ковалевский открыл своеобразный организм - ползающую ктенофору, - названный им Coeloplana metschnikowi. Родовым названием Ковалевский хотел показать, что этот организм объединяет в себе признаки целентерат и планарий, т. е. плоских червей. В 1886 г. другой русский зоолог - А. Коротнев, работая на о. Яве, открыл другую подобную форму, которую назвал Ctenoplana kowalewskii, также указывая в названии объединение в этом организме особенностей ктенофоры и планарий.
В настоящее время в морях Юго-Восточной Азии описан целый ряд подобных форм, объединенных в группу Platyctenidae (плоских ктенофор), но изучение их показало, что не среди них надо искать предков плоских червей, что это просто ползающие ктенофоры без предковых черт организации плоских червей. Вопрос этот приходится решать иным путем. Возможны два варианта. По одному допущению, ктенофорообразные предки первоначально были ориентированы ротовым полюсом ко дну, а аборальным вверх. Затем они испытали расплющивание по основной оси тела и сближение орального полюса с аборальным. В дальнейшем аборальный орган чувств, зачаток мозгового скопления, должен был сместиться на тот участок сплюснутого тела, который стал передним по направлению движения. Таким образом выработались спинная и брюшная поверхности, а ротовое отверстие, как и у многих турбеллярий, осталось в средней части брюшной поверхности. Однако допущение такого пути формирования тела плоских червей должно уступить место другому. Гораздо вероятнее допустить, что ктенофорообразные предки червей ориентировались к дну боком, в таком случае у них сразу формировалась передняя по движению сторона тела, а ротовое отверстие должно было несколько сместиться по брюшной стороне вперед. Такое допущение больше соответствует расположению нервной системы турбеллярий.
Что касается строения тела турбеллярий, то они сохраняют ряд черт радиальной симметрии предков, особенно в строении нервной системы. Они сохраняют также мерцательный эпителий на поверхности тела, расположение рта на брюшной стороне и ряд других особенностей, заимствованных от целентератных предков. Пока не сформировались рычажные конечности, основным механизмом движения оставалось изгибательное движение (рис. 4). Этот тип движения возможен при достаточно мощной мускулатуре и определенном ее расположении "пластами" вдоль всего тела. Оба эти условия сочетаются в кожно-мускульном мешке червей. При этом двигательная мускулатура составляет около половины общего объема тела, а иногда (немертины, пиявки) я значительно больше.

Рис. 4. Различные формы движения у животных: 1 - движение трипаносомы при помощи ундулирующей мембраны; 2 - изгибательное движение полихеты и 3 - рыбы
С образованием конечностей кожно-мускульный мешок распадается на отдельные мышцы. Морфологическая основа двигательной функции кожно-мускульного мешка - расположение сократительных волокон во взаимно перпендикулярном направлении. Это пласты кольцевых и продольных мышц. Еще у грегарин миофибриллы образуют систему продольных и поперечных нитей. Отростки эпителиально-мышечных клеток низших кишечнополостных также образуют слой продольных (от эктодермы) и слой кольцевых сократительных волокон. Однако у кишечнополостных количество мускулатуры невелико, кожно-мускульный мешок не формируется и движение осуществляется реактивным способом - только у ктенофор сохраняется ресничное движение, дающее, однако, при крупных размерах ктенофор очень слабый эффект. Изгибательное, волнообразное движение - весьма целесообразная форма передвижения в водной среде, однако в плотной среде грунтов, особенно морских, эта форма движения не эффективна: у животных возникает гидравлическое прямолинейное движение. При этом формируется обширная полость тела, наполненная полостной жидкостью. Количество мускулатуры в кожно-мускульном мешке уменьшается, но оно достаточно, чтобы сокращением кольцевой мускулатуры тела и перекачкой полостной жидкости вперед обеспечить продавливание хода в грунте, а затем, расширив передний конец тела и заклинив его в ходе, подтянуть заднюю часть тела сокращением продольной мускулатуры.
Принципиально тот же способ движения свойствен двустворчатым моллюскам, пробивающим ход в грунте клиновидной ногой, способной к расширению при перекачке в ее лакуны полостной жидкости, с последующим подтягиванием тела и одевающей его раковины. Интересно отметить, что отличные пловцы - головоногие моллюски - освоили реактивное движение и им не свойственно изгибательное движение тела. У их предков кожно-мускульный мешок уже распался (как и у остальных моллюсков) и была утрачена основа для создания изгибательного движения.
В водной среде возникли и членистоногие, и свойственное им движение с помощью рычажных конечностей, но, прежде чем сказать о свойственной им форме движения, следует остановиться на метамерии (сегментации) и ее происхождении.
Метамерия иногда рассматривается как своеобразный тип симметрии. При го-мономной сегментации, так же как и при других типах симметрии, имеет место повторение сходственных частей тела -метамер (илисомитов) с одинаковым расположением половой системы, выделительных органов, разветвлений нервной (невросомит) и кровеносной (ангиосомит) системы, с двумя параподиями по бокам тела, с поперечными перегородками -диссепиментами, отгораживающими сегменты друг от друга, с обособленными участками продольной и кольцевой мускулатуры (миосомит). Только в данном случае сходственные части -метамеры - расположены не вокруг точки или линии и не по обе стороны плоскости, а в линейном направлении по главной оси тела. Метамерное строение появилось вместе с развитием вторичной полости тела и кровеносной системы (рис. 5, 6). Низшие черви- сколециды - лишены настоящей метамерии (Amera), она появляется только у высших червей - аннелид - и проявляется либо в малом числе сегментов (Oligomera - мшанки, плеченогие и все вторичноротые), либо в большом (Роlymera - аннелиды и членистоногие).
Среди основных этапов развития животного мира наряду с рассмотренными выше - возникновением многоклеточности, радиальных типов симметрии, появлением двусторонней (билатеральной) симметрии - развитие метамерности занимает важное место. Наряду с ними можно поставить еще только формирование рычажных конечностей как основного органа движения и выход из водной среды обитания в воздушную.
Конечности членистоногих и позвоночных возникли совершенно независимо, , хотя они и обладали поразительным биолого-эволюционным и функциональным сходством при резко выраженных чертах различий как следствии различия в характере скелета тех и других. У, членистоногих хитинизированный трубчатый скелет наружный и по расположению и по происхождению (производное эктодермы), у позвоночных скелет костный и внутренний (мезодермический). Функциональное сходство обслуживающей конечности мускулатуры-в антагонистической системе мышц сгибателей и разгибателей, но у членистоногих они прикрепляются к выростам хитинового панциря, а у позвоночных - к выростам костей.
Что же могло вызвать развитие сегментированного тела? Следует прежде всего отметить, что подразделение тела на сходственные части с линейным их расположением наблюдается у разных организмов, стоящих на более низком уровне, чем животные с истинной метамерией. Такие случаи (псевдометамерия) наблюдаются у раковинок фораминифер (рис. 5), турбеллярий, немертин. В этом сказывается упорядочение расположения множественных образований и органов, имеющих полимерный характер.

Рис. 5. Примеры псевдометамерии: 1- 2 раковинки фордминифер; 3 - немертина
Истинная метамерия, появляющаяся у кольчатых червей, - гораздо более общее и глубокое явление, наложившее свою печать на большой и важный этап эволюции полимерных животных - аннелид и членистоногих.
Несегментированные, низшие черви (Amera) немногочисленны, имеют слабые способы передвижения и не имеют в водной среде такого значения, как аннелиды. Наиболее существенный для всех них фактор биотической среды - враги, для которых они могут являться пищей. Турбеллярии ведут придонный образ жизни, малы по размерам, очень медленно движутся, прячутся между водорослями и под камнями. Тело их выделяет много слизи, вероятно, выполняющей защитную функцию. Нематоды ведут скрытый образ жизни - они наполовину паразиты, наполовину живут в грунте и малодоступны для врагов. Среди них нет пелагических форм, и они очень малоподвижны. Такой же образ жизни свойствен приапулидам - они живут в скрытых биотопах, главным образом в грунте, и либо лишены органов движения, либо способны лишь к самому слабому передвижению. К ним относятся коловратки, но они имеют микроскопические размеры и распространены преимущественно в пресных водах. Это единственная группа среди сколецид, ведущая планктонное существование.
Единственная группа среди низших, несегментированных червей, ведущая и придонное и пелагическое существование, многочисленная в морских бассейнах - немертины - агрессивные хищники, обладающие органом нападения и защиты - хоботом. Они движутся волнообразно, изгибая тело, но движение это очень слабо. Кожа их выделяет огромное количество слизи, назначение которой, возможно, двойное: она отпугивает врагов и ускоряет движение в воде. Если не считать еще нескольких немногочисленных групп микроскопических организмов, ведущих донную жизнь, то это и все, что мы находим в морской воде из низших червей. Конечно, самое большое внимание привлекают среди них немертины. Однако и они не смогли занять среди населения моря видного положения.
Для большинства перечисленных групп сколецид характерно отсутствие способности к быстрому движению и плотных, защищающих тело покровов. Для дальнейшей эволюции животных необходимо было разрешить эти затруднения. Некоторые организмы, генетически связанные с червями, находили защиту от врагов в формировании крепких покровов ( плеченогие, мшанки, моллюски), но теряли в скорости движения или даже становились неподвижными, прикрепленными формами. И только многощетинковые кольчецы получили дальнейшее филогенетическое развитие: у них выработались уплотненные, но тонкие покровы и сегментированное строение, которое давало возможность легко сгибать тело во все стороны. Очевидно, первые этапы формирования метамерности были связаны с формированием только поверхностной сегментации, но постепенно она захватывала все более и более глубоко расположенные части тела и органы. Полимеризация захватила некоторые системы внешних и внутренних органов - параподии, жаберные выросты, половую, нервную, кровеносную и выделительную системы. Внутреннее упорядочение в расположении органов подчинилось метамерному строению, и в результате развившейся метамерии возникла гомономная* множественность посегментного расположения систем органов. Так необходимость выработки быстрого движения и защитных покровов нашла свое выражение в метамерном строении.
* (Гомономный - однообразный, от греческих слов homos - одинаковый и nomos -поделенный.)
Очень часто на протяжении эволюции отдельных групп, особенно среди кишечнополостных и червей, наблюдается множественное повторение одноименных образований и органов. Такие структуры именуютполимерными. Особенно часто встречается полимерный тип в расположении половых желез. Их четыре у гидрозоа и сцифозоа, много у антеозоа, 8 мужских и 8 женских желез у гермафродитных ктенофор, очень много у ленточных червей ( цестод), много у полихет.
Целый ряд полимерных органов, помимо гонад с посегментным расположением, возник при формировании метамерных кольчецов - параподии, жабры, выделительные органы, кольцевые нервные тяжи и кровеносные сосуды, ганглиозные скопления в брюшной нервной системе и многое другое. Полимерный характер имеет расположение многих органов у иглокожих в связи с их пятилучевой симметрией. Подобных примеров можно было бы привести очень много и для членистоногих - количество ног у ракообразных и особенно у многоножек, трахеальные образования у них же и у насекомых и т. д. Однако в дальнейшем почти всегда наблюдается процесс уменьшения количества одноименных образований, или, как это называют, процессолигомеризации, в количестве ног, сегментов тела, половых желез, органов выделения, дыхания и т. д.
Гомономная сегментация стала уступать местогетерономной* и особенно в группе членистоногих, а покровы становились все более прочными (хитинизированными), но не утерявшими своей легкости и не мешавшими подвижности благодаря мягким и растяжимым сочленениям между отдельными более плотными частями хитинового панциря. В дальнейшем в эволюции обеих основных ветвей животных - первичноротых и вторичноротых - проявляется поразительное сходство в выработке рычажных двигательных придатков - конечностей. У членистоногих прообразом их были параподии полихет, у позвоночных - плавники рыб. У членистоногих конечности появились еще в водной среде (у ракообразных), а у позвоночных только с выходом в воздушную среду. Эволюция параподии и конечностей у полихет и членистоногих - очень хороший пример олигомеризации и перехода от гомономных образований к гетерономным. У многих полихет параподии имеются на всех сегментах и иногда на сотнях сегментов имеют совершенно одинаковый характер (гомономная полимерия). У ракообразных и тем более у паукообразных и насекомых количество конечностей подвергается неуклонно идущей олигомеризации и гетерономному типу изменений, т.е. уменьшается не только количество ног, но и разные конечности отличаются друг от друга по строению и функциям. Этот процесс представляет собой только часть более общих изменений организации процессакефализации** (формирование головы) итегматизации*** (формирование отделов тела). Из передних пяти-шести сегментов тела формируется голова - передняя тегма тела, а конечности этих сегментов превращаются в органы чувств и ротовые придатки. Следующие сегменты образуют грудной отдел (у насекомых 3 сегмента, у паукообразных - 4, у высших ракообразных - 8 и т. д.).
* (Гетерономный - различно поделенный, от греческого heteros - неодинаковый.)
** (Kephalon (греч.) - голова.)
*** (Tegma (греч.) - отдел.)
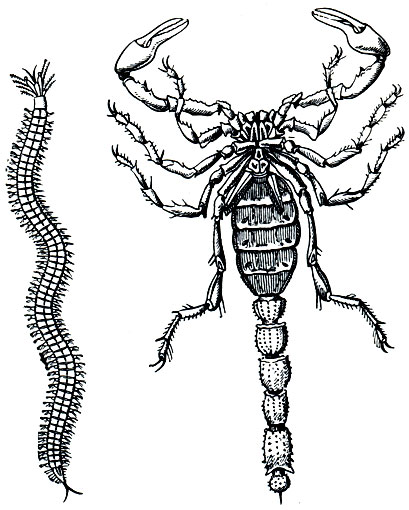
Рис. 6. Примеры истинной метамерии: слева - гомономная (полихета); справа-гетерономная (скорпион)
Конечности грудного отдела обычно служат для передвижения. Брюшной отдел, сформированный из различного количества сегментов, либо лишен конечностей (насекомые, паукообразные), либо они выполняют другую функцию (половую, дыхательную и т. д.). Грудь и брюшко образуют вторую и третью тегмы тела.
У наземных групп конечности выполняют основную двигательную функцию (если не считать многоножек, змей и некоторых других групп), а сегментация как во внешнем облике животного, так и во внутреннем строении исчезает.
Метамерное расположение сохраняется лишь частично и только на некоторых системах, таких, как дыхательная, нервная и др. Крайнюю форму подавления (исчезновения) метамерного строения можно наблюдать у клещей, в мешкообразном теле которых слиты все отделы, не обнаруживающие сегментированного плана строения. И у других членистоногих намечается слияние трех основных отделов тела, прежде всего головы и груди в головогрудь (высшие ракообразные и паукообразные). При этом иногда брюшной отдел тоже испытывает редукцию, и тогда, как, например, у крабов, тело кажется состоящим из одного компактного образования, хорошо защищенного крепким панцирем от врагов, а хорошая подвижность его обеспечивается пятью парами конечностей.
Столь же нагляден этот процесс и у позвоночных. Две пары конечностей, сходные между собой у низших позвоночных, особенно у рыб, у птиц совершенно различны - передние служат для полета, задние для передвижения по грунту. У многих млекопитающих строение и функции передних и задних конечностей также весьма различны. Здесь процесс олигомеризации выражен очень ясно.