

Симбиоз, комменсализм и паразитизм
* (Симбиоз - совместное существование, от греческих слов sym - вместе и bios - жизнь; комменсализм - нахлебничество, от латинских слов cum - с и mensa - стол; паразитизм от греческих слов para - около и sitos - пища.)
Чрезвычайно многообразны взаимоотношения, которые складываются между организмами и их группами, а иногда и целыми биоценозами в длительном процессе эволюции приспособлений и биологических связей.
Об одной такой системе связей между растительным и животным царствами в рамках всей биосферы мы уже говорили.
Планктонные биоценозы дают основной пищевой ресурс для донной фауны океанов и морей, а также и для рыб. Многие организмы моря и суши совершают горизонтальные, а в море, кроме того, и вертикальные миграции и на разных этапах миграций вступают в биологические связи с различными группами организмов.
Многие паразиты часть своей жизни проводят в свободном состоянии, и тогда они подчиняются иным системам связей с другими организмами и с абиотической средой.
Одна из ведущих связей между организмами - это потребность в пище. В этом отношении животные в первую очередь могут быть разделены по характеру пищевого объекта на растительноядных, детритоядных, грунтоядных, плотоядных, в том числе и хищников.
Понятие "хищник" употребляется в различном смысле.
Животных, питающихся моллюсками, насекомыми или червями, обычно не называют хищниками, но совершенно очевидно, что треска, питающаяся сельдью, - хищник, так же как акула, пожирающая других рыб, или волк, гоняющийся за зайцем. Обычно хищниками называют животных, питающихся другими животными, близкими по систематическому положению (в рамках позвоночных, или насекомых, или моллюсков и т. д.), но иногда говорят о хищных растениях, питающихся насекомыми, как, например, росянка, или о хищных грибах, питающихся червями, и т. п. Более точное определение и классификация способов питания, вероятно, затруднительны в силу того, что эти способы создаются независимо у разных животных в совершенно разных формах, на различной морфологической основе и не имеют никакой генетической связи. В силу этого создание естественной системы форм питания едва ли возможно. Система всегда будет носить более или менее искусственный характер. Она возможна, пожалуй, только в рамках небольших однородных групп.
Животных можно различать и по способу захвата пищи -фильтраторы* ( губки, двустворчатые моллюски, усоногие ракообразные, усатые киты и др.),грунтоеды и различные группы животных, обладающих в питании избирательной способностью,-зерноядные инасекомоядные птицы,хищники и т. д.
* (Фильтраторы-животные, пропускающие через особые полости большое количество воды и отфильтровывающие при этом планктон.)
Часто среди животных, входящих в один морской донный биоценоз, наблюдается мозаичность в использовании пищевых ресурсов: фильтраторы могут жить рядом с грунтоедами и собирающими, а иногда они специализируются по отдельным кормовым объектам. Обычно именно таким образом подбираются по способу питания животные в биоценозе. Так, в Каспийском море осетр питается почти целиком моллюском синдесмией, а севрюга почти нацело - полихетой нереис. Оба эти организма перед второй мировой войной были перенесены в Каспий из Азовского моря и получили в нем огромное развитие.
По объектам питания животных можно также разделить намонофагов иполифагов (т. е. питаются одним объектом или многими).
В системе биологических связей между животными организмами можно выделить несколько типов с постепенным усилением биологической связи -комменсализм,симбиоз ипаразитизм.
Комменсализм, илинахлебничество,- очень обычное явление у общественных насекомых. Особенно обильны комменсалы в гнездах муравьев, где они существуют за счет запасов, накопляемых хозяевами гнезда. Среди них есть и несколько видов жуков семейств Staphylinidae, Pselaphinidae и др. Типичные комменсалы - рыбы-лоцманыж рыбы-прилипалы, сопутствующие акулам, жуки в гнездах муравьев и т. п.
Чрезвычайно разнообразно явлениесимбиоза, который может быть двусторонне полезным (мутуализм*), односторонне полезным или двусторонне нейтральным. Классический пример двусторонне полезного симбиоза дают рак-отшельник и актиния, сидящая на его раковине. Актиния пользуется остатками со стола своего хозяина, который, кроме того, является для нее извозчиком, а рак-отшельник (Eunagurus) имеет защитника - актинию, отпугивающую его врагов. Рак-отшельник активно разыскивает актиний, снимает их клешней с субстрата, на котором они сидят, и пересаживает к себе на раковину, в которой живет.
* (Мутуализм-двусторонне полезный симбиоз, от латинского слова mutuus - взаимный.)
Обычны различные формы симбиоза между некоторыми морскими животными и одноклеточными водорослями - так называемыми зоохлореллами (зелеными) и зооксантеллами (желтыми). Зооксантеллами набита протоплазма некоторых радиолярий, ими набиты ткани некоторых кораллов, причем если кораллы освободить от зооксантелл, то нормальная жизнедеятельность кораллов нарушается. Недавно было показано, что зооксантеллы - это молодые стадии развития одноклеточных водорослей перидиней.
Самый огромный моллюск Tridacna, достигающий размера в 11/2 м и веса в 200- 300 кг, дает приют огромной массе одноклеточных водорослей, без которых существовать не может. Турбеллярия Convoluta, развивающаяся в огромном количестве на литорали Англии и Нормандии, имеет ярко-зеленую окраску из-за набивающей ее ткани одноклеточной водоросли Carteria (зоохлорелла). Это также двусторонне полезный симбиоз: животное защищает водоросли, дает им углекислоту и азотистые соединения, а само получает от водорослей кислород и частично использует их как пищу. Известно, что если молодая конволюта не получит порцию водорослей для дальнейшего развития, она погибает.
Различного типа симбиозы, правда, не столь резко выраженные, весьма многочисленны. Стайки молодых рыб находят приют среди щупалец некоторых медуз, что является примером односторонне полезного симбиоза. В заднем отделе кишки голотурии Holothuria tubulosa обитает маленькая рыбка Teraster umberis; она выплывает из голотурии, но при опасности мгновенно скрывается в своем убежище.
Совсем особая группа биологических связей между организмами - явление паразитизма. Паразитизм очень широко распространен среди многих групп животных. Особенно широко паразитизм распространен среди простейших, червей и членистоногих (цв. табл. I).
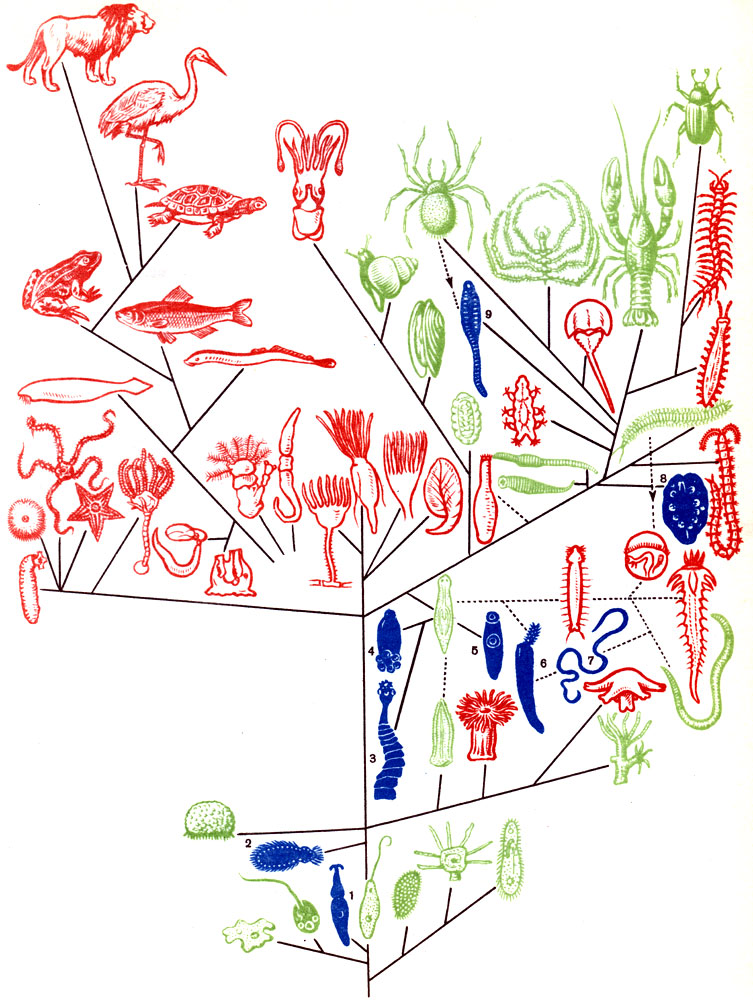
Таблица 1. Распространение паразитизма в разных группах животных. На генеалогическомдреве синим цветом изображены классы, целиком состоящие из паразитов, зеленым - классы, содержащие наряду со свободно живущими, также и паразитические формы, и красным - классы, образованные исключительно свободноживущими видами. 1 - Gregarina; 2 - Mesozoa; 3 - Cestodes; 4- Monogenea; 5 - Digenea; 6 - Acanthocephala; 7- Nematodes; 8 - Myzostomida; 9 - Pentastomida
Имеется немало классов животных, ведущих исключительно паразитическое существование. К ним относятся споровики, трематоды, цестоды, колючеголовые и некоторые другие черви, из ракообразных - мешкогрудые (Ascothoracida). С другой стороны, ни одна группа животных не свободна от паразитов и симбионтов. Существует очень много попыток дать общее определение и классификацию явлений паразитизма, но удачного решения этой задачи пока не найдено. Явления паразитизма возникали в животном царстве много раз и независимо друг от друга. В силу крайнего многообразия явлений паразитизма и множественности путей возникновения их классификация имеет искусственный характер.
Определение паразитизма включает два положения: 1) организм хозяина служит средой обитания (паразит обитает на теле хозяина или внутри него) для паразита, и внешняя среда по отношению к телу хозяина воспринимается паразитом опосредствованно через тело хозяина, 2) паразит использует хозяина как источник питания, он питается тканями хозяина или переваренной им пищей.
Естественно задать вопрос: каким же образом возникает паразитическое существование? Что заставляет свободно живущий организм переходить к паразитизму и существованию внутри другого организма, казалось бы, в очень сложных и трудных условиях? Этот вопрос аналогичен такому же вопросу в отношении глубоководной фауны, фауны пещер и почв. Найти правильный ответ на этот вопрос мы можем, только учитывая способность живых существ к расселению и освоению новых биотопов и новых биологических особенностей. Это свойство настолько резко выражено, что многие организмы путем длительного процесса приспособления заселяют чуждые им аномальные биотопы, соответственно которым изменяется их организация. Освоение этих биотопов при переходе к паразитическому существованию идет обычно несколькими каналами. Очень часто паразит проникает в тело хозяина, внедряясь в его пищеварительную систему в личиночном состоянии вместе с пищей. Бесконечно повторяющаяся инвазия может кончаться успехом, и внедряющийся организм вырабатывает способность существовать, а затем питаться, расти и размножаться в кишечнике другого животного. К паразитическому существованию может приводить хищничество, если хищник очень мелкий по сравнению с жертвой - хозяином. Он может сам все теснее и теснее вступать в связь с поверхностью тела хозяина и в конце концов проникать в него. Кровососущие животные ( мухи, комары, клопы, клещи) могут при этом заносить в тело жертвы паразитов. Часто на теле крупных животных поселяются мелкие животные, которые становятся их постоянными спутниками, а в дальнейшем также проникают под покровы их тела во внутренние органы.
Существуют, конечно, и другие пути формирования паразитизма. Вероятно, путем постепенного приспособления к хозяину к паразитизму могут переходить комменсалы и симбионты.
Явления паразитизма связаны с массой замечательных и своеобразных особенностей. Нервная система и органы чувств, а также мускулатура и органы движения обычно испытывают у паразитов явления регресса. В силу всех этих изменений иногда взрослый паразит теряет всякое сходство с родственными группами, как, например, у некоторых брюхоногих моллюсков (Parenteroxenos) или ракообразных (из веслоногих и усоногих, в том числе знаменитая Sacculina).
Сильное развитие приобретают органы прикрепления к телу хозяина (крючки, присоски, хоботки) и защитные от хозяина образования покровов тела, но наиболее характерное отличие - в сильном и часто в колоссальном развитии плодовитости. Нередко из одного оплодотворенного яйца образуются (партеногенетически) тысячи, сотни тысяч и миллионы особей. Количество яиц, продуцируемых одной самкой турбеллярий (свободноживущих плоских червей), исчисляется единицами или десятками, а у близкородственных им трематод - многими тысячами и даже десятками тысяч. Интересно при этом отметить, что у родственных между собою трематод и цестод имеются еще дополнительные механизмы к увеличению числа потомства, при этом совершенно разные у двух несомненно родственных форм. У ленточных червей увеличивается число половых систем, возникающих почкованием, причем их количество в одном черве может достигать нескольких тысяч. Совершенно иначе увеличивается плодовитость трематод - в спороцистах и редиях (см. раздел о трематодах) происходит партеногенетическое размножение яиц (шары дробления) на ранних стадиях развития. Часто организм ведет паразитическое существование только в определенные периоды своего жизненного цикла: или в раннем личиночном возрасте, или во взрослом состоянии. В остальное время он существует во внешней среде как свободноживущий организм (различные нематоды). В других, довольно многочисленных случаях паразит оказывается на свободе только в стадии яйца ( трематоды, цестоды и нематоды) и, наконец, может совсем не существовать в свободном состоянии ( малярийный паразит, трихина и др.). Усложнение биологии паразита возникает в связи со сменой хозяев: паразит проводит часть цикла в одном хозяине, часть в другом.
При этом основным хозяином считают организм, в котором женские гаметы, или яйца, проходят оплодотворение, а другого хозяина называют промежуточным. В наиболее сложных циклах промежуточных хозяев может быть два, например у широкого лентеца. Первый промежуточный хозяин - пресноводный рачок циклоп, а второй - рыба. Своеобразное усложнение паразитического жизненного цикла у споровиков заключается в множественной повторности бесполого размножения (шизогония*).
* (Шизогония - множественное деление; от греческих слов schizein - расщепляться и goneia - воспроизводство.)
Очень интересные формысверхпаразитизма наблюдаются у насекомых (мухи и перепончатокрылые): насекомое откладывает яйца в паразитическую личинку другого насекомого, находящуюся внутри личинки третьего.
Не менее интересен вопрос специфичности хозяина в явлениях паразитизма. Некоторые паразиты могут существовать в теле только одного или двух хозяев. Так, некоторые виды споровиков рода Plasmodium могут развиваться только в теле комара Anopheles, где проходят половой цикл, и в теле человека, где протекает бесполое развитие (шизогония). Возбудитель сонной болезни - Trypanosoma gambiense - нормально обитает в крови антилопы, для которой, видимо, не патогенен, переносчиком его является муха цеце (Glossina morsitans), а человек - дополнительным, случайным носителем. Свиной солитер может развиваться только в теле свиньи (промежуточный хозяин) и человека (основной хозяин). Наконец, часто хозяин не специфичен и паразит может развиваться в различных животных. Так, эхинококк может и во взрослой стадии, и в стадии финки существовать в различных животных, в том числе и в человеке. Имеются паразиты с еще более широким кругом хозяев.
Весьма замечательны, хотя еще мало изучены, явления физиологических и биохимических связей между хозяином и паразитом. Разве не замечательно, что при нормально протекающей малярии совпадают циклы шизогонии паразита, сидящего в бесчисленных эритроцитах крови хозяина (наступление пароксизма), и что микрофилярии появляются в периферических кровеносных сосудах в ночные часы, когда на человека нападают кровососущие насекомые, переносящие микрофилярии на другого хозяина (см. в соответствующих разделах книги)!
Явления паразитизма, симбиоза и комменсализма - интереснейший раздел биологии, имеющий большую перспективу дальнейшего изучения, полный глубокого научного смысла и огромного практического значения.
Стадии покоя, спячки и анабиоза у животных. Животные в состоянии нормальной жизнедеятельности могут существовать в различных условиях. В арктических морях существует немало форм, вся жизнь и размножение которых протекают при температуре ниже нуля, а иногда 1,5- 1,8°С ниже нуля, например моллюск Portlandia arctica, офиуры - Ophiopleura borealis и Gorgonocephalus arcticus, звезды - Asterias panopla и Pontaster tenuispinus, креветка Sclerocrangon ferox, морской таракан Mesidothea sabini и др. С другой стороны, коралловые рифы не могут развиваться при температуре ниже 21,5° С. И те и другие относятся к так называемымстенотермным* организмам.
* (Стенотермный - существующий в условиях слабых колебаний температуры, от греческих слов stenos - узкий и termos - тепло. Противоположный термин-эвритермный, eurys - широкий.)
Множество пресноводных организмов погибает при повышении солености, что и происходит в течение миллионов лет в устьях рек при изливании их вод в моря. Но вместе с тем в пересоленных (гипергалинных)* озерах при почти предельном насыщении солями существуют и развиваются, да еще в огромных количествах, листоногий рачок Artemia salina и служащее ему пищей жгутиковое Dunaliella. С другой стороны, существует фауна горячих источников - это животные, жизнедеятельность которых может протекать при температуре до 40-50° ( нематоды, личинки мух, брюхоногие моллюски).
* (Гипергалинные - существующие в условиях повышенной солености; hiper - сверх, halos (греч.) - соленый. Противоположный термин - олигогалинный; oligo (греч.) - мало.)
То же самое можно сказать и о способности животных переносить пониженное количество кислорода. Громадное большинство животных может существовать только при достаточном насыщении среды кислородом. Имеются и такие, которые не выносят даже слабого понижения в содержании кислорода. Например, турбеллярия Planaria alpina или ракообразное Bythotrephes -стенооксибионтные* организмы. Однако имеются животные из разных групп, которые могут существовать при очень малом количестве кислорода в окружающей среде или совсем без него, как некоторые водные личинки двукрылых, некоторые низшие ракообразные, нематоды и др.
* (Стенооксибионтные - требующие для существования строго определенного снабжения кислородом, от греческих слов stenos - узкий, oxys - кислый, bios - жизнь. Противоположный термин - эвриоксибионтный, от eurys - широкий.)
Среди приспособлений, при помощи которых животные переживают неблагоприятные для них условия существования, видное место занимает временный переход в состояние покоя (латентное состояние). Все животные в большей или меньшей степени способны в течение очень короткого срока переносить неблагоприятные условия без специальных приспособлений. Правда, клещи, клопы и некоторые другие кровососущие животные могут обходиться без пищи в течение очень длительного времени, даже нескольких лет, но это редкое исключение. Способность сохранять жизнь при наступлении неблагоприятных условий значительно расширяется переходом в не жизнедеятельное состояние, в стадию покоя - зимующие яйца, цисты, споры, спячка, анабиоз. Формирование этих образований может наступать или при изменении условий среды в неблагоприятную сторону, или связано у паразитических форм с выходом во внешнюю среду. В северных и умеренных широтах многие животные впадают в спячку в зимнее время, а в жарких и сухих странах - в период летней засухи.
Зимние, или покоящиеся, яйца свойственны очень многим пресноводным микроскопическим животным - коловраткам, ластоногим и ветвистоусым ракообразным, которые способны переносить высыхание окружающей среды и низкую (зимнюю) температуру. Яйца глистов также могут длительное время находиться во внешней среде в покоящемся состоянии. Яйца рачка Artemia salina сохраняют жизнеспособность в течение 9 лет. Сходного типа образования имеются у пресноводных губок и мшанок (геммулыи склеробласты), но это не яйца, а многоклеточные комплексы (см. соответствующие разделы книги).
Иного типа образования -споры ицисты. Первые свойственны споровикам и входят в их цикл как определенная стадия развития, связанная обычно с выходом во внешнюю среду, и поэтому у кровяных споровиков, которые никогда не оказываются во внешней среде, стадия споры утрачивается.
Многие пресноводные простейшие ( корненожки и инфузории) и микроскопические многоклеточные ( тихоходки, нематоды, коловратки и некоторые другие) в случае наступления неблагоприятных
условий способны выделять оболочки и переходить на длительное время в состояние покоя. Нередко при этом можно различить две оболочки цисты -эндоцисту, очень тонкую и обладающую большой химической стойкостью, иэкзоцисту, обладающую механической прочностью.
При образовании цист у простейших наблюдаются существенные изменения в протоплазме - выделяется большое количество воды, протоплазма густеет, теряются реснички и т. п. Видимо, такого же характера изменения происходят и при образовании любых стадий покоя. Эти изменения дают организмам возможность переносить сильное повышение и понижение температуры, высыхание, понижение количества кислорода. Мелкие нематоды и тихоходки (Tardigrada) в состоянии цист могут переносить временный нагрев до 150° С выше нуля и охлаждение до 270° С ниже нуля.
Споры многих бактерий, коловраток, тихоходок и некоторых нематод в сильно высушенном состоянии могут переносить охлаждение до температуры жидкого воздуха и даже до-250° С. Многие насекомые, млекопитающие и другие животные могут переносить в состоянии переохлаждения (но не замерзания) температуру ниже нуля: у сусликов до -0,2°С, у летучих мышей до 1 - 2° С и т. п., не погибая даже при длительном пребывании в этой температуре. Все описанные стадии покоя имеют и другое биологическое значение: в виде спор и цист простейшие в высушенном состоянии переносятся на большие расстояния и таким образом расселяются. Поэтому среди образующих цисты простейших наблюдается нередкокосмополитизм*.
* (Космополитизмом в биологии называют очень широкое распространение в пределах своего основного биотопа, от греческих слов cosmos - мир и polyteien - гражданин.)
Явление перехода в недеятельное состояние (спячка,анабиоз) известно и у многих выше стоящих по уровню развития организмов - у высших червей, моллюсков, насекомых и у всех классов позвоночных - без образования спор и цист, причем степень перехода в покоящееся состояние (спячки, как это иногда называют) бывает различна. Для растений и некоторых животных переход в такое состояние дает основание говорить о полной приостановке жизненных процессов, но мнения ученых по этому вопросу расходятся: одни считают, что полный перерыв жизненного обмена невозможен, другие считают, что возможен. Какие же явления сопровождают переход в спячку? Прежде всего потеря огромного количества воды иногда почти до полного ее отсутствия. Дождевые черви и наземные моллюски могут при высыхании терять до 80% и более воды и сохранять способность к "оживанию". Наиболее способные к высыханию земноводные и пресмыкающиеся теряют до 50% воды.
Во время спячки все процессы жизнедеятельности организма замедляются (дыхание, выделение) или даже приостанавливаются (питание). Ранее предполагали, что при полном промерзании организма ( лягушки, рыбы, насекомые) он может оживать, как часы с остановленным маятником, если он вновь пущен. Однако крупные специалисты в этом вопросе П. Ю. Шмидт и Н. И. Калабухов утверждают, что полное замерзание жидкостей тела всегда убивает организм.
Переселения и акклиматизация. Каждый организм характеризуется определеннымареалом*, в границах которого он находит необходимые для него условия существования и размножения. Внутри ареала организмы большей частью распространены неравномерно, в некоторых его участках достигая наибольшего развития, а в других - среднего, или встречаются спорадически. В связи с этим можно различать распространение целостное или разорванное. Последнее обычно характерно для краевых зон ареала и для реликтовых форм. Подреликтами** подразумевают формы, основной ареал которых расположен в другом районе, в условиях, которые ранее существовали и в данном месте, но в силу климатических или других изменений ему уже не свойственны. Так, в Московской области мы находим в разных местах, часто на большом расстоянии одно от другого, ледниковые озера, окруженные торфяниками, со всем типичным для торфяника набором растительных и животных форм, мы их называем ледниковыми реликтами. Далеко на севере эти торфяники имеют свой основной ареал, а под Москвой сохраняются как реликты.
* (Ареал от греческого слова area - поле, площадь.)
** (Реликт от латинского слова relinquere - оставаться.)
Реликты могут быть связаны не только с низкой температурой. В Белом море в изобилии произрастает морская трава - зостера. Эта часть ее ареала оторвана от основного ареала, доходящего на севере до западного Мурмана. На восточном Мурмане ее нет, но в литориновое более теплое время несколько тысяч лет назад ареал зостеры был сплошным. Последующее похолодание разорвало ареал зостеры, и она сохранилась в Белом море как тепловодный реликт. Под воздействием изменяющихся условий основной и часто более обширный в прошлом ареал распадается на отдельные реликтовые местонахождения. Таким холодновидным реликтом может считаться в Северной Европе и Сибири равноногое ракообразное - морской таракан (Mesidothea etomon) (кроме того, он еще и солоноватоводный реликт, так как сформировался в водоемах малой солености послеледникового времени). По своему происхождению и распространению сходно с ним другое ракообразное - Mysis relicta. Обе эти формы вместе с рядом других подобных форм можно найти в наиболее опресненных участках окраин наших северных морей, а также в пресных озерах Швеции, Северной Европы и Сибири и даже в Каспийском море. У всех них десяток тысяч лет назад был огромный сплошной ареал, а ныне они остались только в различных участках как арктические солоноватоводные реликты. Уходящий на север тающий ледниковый покров разбросал их в виде реликтов на огромном пространстве Европы и Азии. Таким образом, с понятием реликта обычно связывается существование организма в географических зонах, общие климатические условия которых претерпели значительные изменения. В каком-то смысле реликтами можно считать и различные формы, исчезнувшие, исчезающие или сократившие численность под воздействием человека. Возможно, к ним надо отнести и еще недавно исчезнувших стеллерову корову в морях Дальнего Востока, птицу додо с Маврикиевых островов, лошадь Пржевальского и др., а сейчас относятся зубр, морской котик, морская выдра и многие другие.
Если брать ареалы современных животных, находящихся в условиях процветания, то и среди них мы можем найти формы с очень широкими, почти всесветными ареалами распространения. Их иногда именуютубиквистами*, или космополитами. Абсолютных убиквистов мы, пожалуй, не знаем, обычно они ограничены или водной средой, или воздушной. Многие из них развезены по всему миру человеком. Имеется немало и таких форм, которые имеют небольшие ареалы. Но подавляющее большинство животных имеет ареалы, ограниченные определенными непреодолимыми для них преградами - границами суши и морей, горными хребтами, пустынями, широкими реками и т. п.
* (Убиквисты - от латинского ubique - повсюду, то же, что космополиты.)
Вряд ли можно представить себе такое животное, которое заселяло бы все те биотопы биосферы, которые благоприятны для его существования. Наоборот, в огромном большинстве случаев фактический ареал - это только часть того ареала, в котором данный организм мог бы существовать, если бы непреодолимые для него препятствия не ограничивали его стремления и возможностей к расселению. Надо учитывать и то, что указанные непреодолимые преграды претерпевают в геологическом масштабе существенные изменения.
Английские острова были еще недавно частью Европы, обе Америки соединились только в середине третичного периода, а Азия и Америка разъединились в недавнее послеледниковое время. Мадагаскар в мезозойское время был частью африканского материка.
Еще в третичное время Черное, Азовское и Каспийское моря входили в один морской бассейн и т. д. С другой стороны, существуют и очень древние разрывы частей суши. Еще Уоллес отметил, что глубокий морской пролив между островами Бали и Лембок разорвал пути развития австралийской и азиатской фауны на долгие геологические времена.
Ограниченность ареалов зависит еще и от способности организмов к расселению. Например, некоторые простейшие, особенно пресноводные корненожки, коловратки, тихоходки и некоторые другие, обладающие способностью образовывать цисты и стадии покоя и переносить в этом состоянии неблагоприятные для жизнедеятельных форм условия, в том числе и высыхание, могут разноситься ветром, птицами и разными другими способами на огромные пространства и приобретать всесветные ареалы. Все другие животные, однако, ограничены в своем распространении, и для большинства из них потенциальные ареалы, т. е. части биосферы, где они могут существовать, превышают фактические ареалы, и часто в огромных масштабах. Это подтверждается множеством примеров.
За последние 40 лет наши арктические моря претерпели значительное потепление, и в результате вся населяющая их фауна переместилась с запада на восток: тепловодная фауна - из Северной Атлантики в Баренцево море, из него дальше - в Карское море, а холодноводная фауна, наоборот, отступила из западной и юго-западной частей Баренцева моря на восток и север. Однако гораздо показательнее те случаи, когда животное преодолевает непреодолимые для него преграды, в подавляющем числе случаев с сознательным или бессознательным участием человека.
Каспийскийэндемик* гидроид Соrdylophora caspia, очевидно, с помощью судов, днища которых он обрастает, во второй половине прошлого века по Мариинской системе проник в Балтийское море, а оттуда кораблями был развезен по всем крупнейшим портам мира. Китайский мохнатоногий краб (Eriocheir sinensis) тоже с кораблями перед первой мировой войной проник с Дальнего Востока в устье Эльбы, откуда широко расселился по побережьям Северного и Балтийского морей и речным бассейнам. В новом ареале он достиг огромного развития. Тем же путем брюхо-ногий моллюск рапана (Rарапа besoar) переселился около тридцати лет назад из Японского моря в Черное, размножился в нем и стал злостным вредителем устрицы и мидии и излюбленным сувениром черноморских туристов. Еще один краб, Rhithropanopeus harrisi, совершил более сложное переселение с тремя "пересадками". Сначала от побережий Северной Америки через Атлантический океан в Зюйдерзее, оттуда - к нам в Черное море, а из него Волго-Донским каналом в Каспийское море, и в каждом новом местообитании этот маленький крабик получил массовое развитие. Крабик был завезен также с западного побережья Северной Америки на восточное. Любопытно, что все эти путешествия вплоть до Каспийского моря крабик ритропанопеус совершил с полихетой Marcierella enigma, вместе с которой живет и в исходном ареале. И таких примеров непрошеных "нарушителей границ" мы знаем великое множество - американская виноградная тля филлоксера, колорадский жук, проникший 90 лет назад из Северной Америки в Германию, а сейчас неуклонно стремящийся на восток. Иногда эти пути идут и в обратном направлении.
* (Эндемик - от греческого endemos-местный, живущий только в данном месте. Могут быть эндемики узкие, например озеро Севан, и эндемики широкие, например озера Северной Америки или Европы.)
Развитие авиационных путей сообщения привело к завозу из Северной Америки малярийных комаров в Бразилию. Через несколько лет комары распространились в глубь страны и вызвали жестокую эпидемию малярии. В начале нашего столетия из Европы в Северную Америку был завезен европейский скворец (Sturnus vulgaris), распространившийся к середине столетия в огромном количестве по всей стране. Есть много и других подобного типа примеров.
Понимание того, что организм может существовать не только в пределах его фактического ареала, но и далеко за его пределами, возникло давно, и на нем базируется вся деятельность человека по акклиматизационным мероприятиям. Число таких мероприятий огромно. Начало им было положено переселением в новые места обитания домашних животных и культурных растений. Нынешние флора и фауна Австралии и Новой Зеландии имеют совсем иной характер, чем сто лет назад. В значительной степени изменились флора и фауна Европы и Северной Америки, акклиматизационные мероприятия постепенно захватывают весь земной шар. Из наиболее ярких примеров можно привести рыбку гамбузию (рис. 8) и ондатру (рис. 9). Однако известны случаи, когда очень хорошее по первоначальным результатам акклиматизационное мероприятие давало в дальнейшем и отрицательный результат. Так, рыбка гамбузия настолько размножилась в некоторых водоемах, например, Средней Азии, что выела все корма для других рыб и подорвала их запасы. С безобидным, казалось бы, кроликом в Австралии пришлось вести борьбу, так как огромные его стаи стали угрозой культурным растениям.
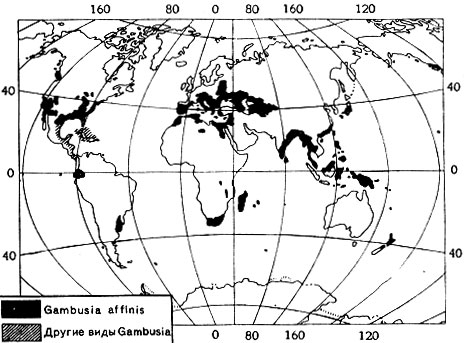
Рис. 8. Распространение североамериканской рыбки гамбузии по земному шару
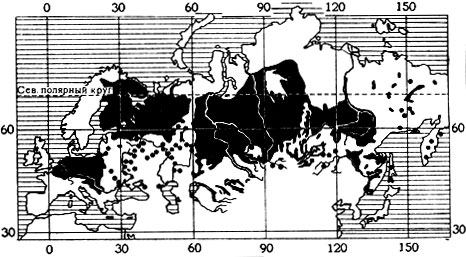
Рис. 9. Распространение американского грызуна ондатры в Европе и Азии
Большое количество подобных фактов описывается в интересной книге английского эколога Элтона "Экология нашествий животных и растений", переведенной и изданной в СССР в 1960 г.
Интереснейшая деталь очень многих успешных акклиматизаций в новом ареале - это исключительная массовость развития, превышающая во много раз степень развития в исходном ареале. Такую массовость развития вновь появившейся в новом ареале формы можно уподобить как бы некоему биологическому взрыву. В 1911 г. из Японии в США был случайно завезен майский жук Popillia japonica, достигший в новом местообитании колоссального развития. Уже через 8 лет в местах его массового развития один сборщик мог собрать в течение дня до 20 000 жуков.
В течение последних 40 лет яркие примеры таких биологических взрывов дают многие новые "вселенцы" в Каспийское море. Во время гражданской войны случайно в Каспий попал из Черного моря двустворчатый моллюск Mytilaster lineatus, ставший вскоре в этом море самым массовым. В тридцатые годы такого же массового развития достигли два вида креветок рода Leander. Перед второй мировой войной в Каспийское море были перенесены из Азовского полихета Nereis diversicolor и двустворчатый моллюск Syndesmya ovata для обеспечения пищей осетровых рыб. Последующее развитие обеих форм в Каспии также оказалось подобным биологическому взрыву, и их общее количество измеряется многими миллионами центнеров. С прорытием Волго-Донского канала переселение из Азовского моря в Каспийское различных растений и животных приобрело стихийный характер, и в настоящее время "вселенцы" составляют в Каспии гораздо большую массу, чем своя исконная фауна. Было несколько удивительно, что балтийская сельдь салака дала в Аральском море, куда ее пересадили, крупную по размерам, вполне жизнеспособную расу. Исключительный интерес представляет успешная акклиматизация дальневосточных лососей в водах Новой Зеландии и Южной Америки.
Все изложенное хорошо поясняет, какой огромной важности оружие приобретает человек для своего хозяйства, используя способность многих растений и животных давать массовое развитие в новом ареале, и сколь необходима теоретическая разработка проблемы акклиматизации для наиболее разумного использования этой способности.